В страните от средиземноморска Европа гилдията на зидарите приличала на другите занаятчийски гилдии и съществувала за благото на каменоделците и понякога и на архитектите. Но днес Братството на свободните зидари (масони) обединява членове от всички области на живота, посветили се на „изповядване на морални и социални добродетели“. В тази си форма то е създадено в Англия през четирийсетте години на XVII век от един литературен кръг, събран около Елиът Ашмол (1617-1692), по-късно спонсор на колекцията на първия английски музей, „Ашмолиън“. Имитирайки средновековната гилдия, те приемат редица графични символи, между които компаса и Слънцето, и определят правила за взаимопомощ. Смята се, че символите са заимствани от тамплиерите и розенкройцерите.
Символ на свободните зидари
Розенкройцерство
Розенкройцерите са тайно общество и е трудно да се открият подробности за неговото начало. Те твърдят, че са последователи на германския окултист и химик Кристиян Розенкройц, живял през XIV или XV в. след Хр. Първата им поява пред външния свят е през 1614 и 1615 г. с публикуването в Касел, Германия, на някои книги, между които „Fama Fraternitalis“ („Братство на сияйния Орден на розовия кръст“), 1614г. Те тълкуват мистични знаци и работят с алхимични формули.
Германски археолози са открили бойни полета от периода на ранната бронзова епоха.
Сериозно наранени и счупени човешки останки, датиращи от ранната бронзова епоха, са били открити по брега на немска река, недалеч от гр. Нойбранденбург. Това откритие доказва, че в онова далечно време на това място се е провело голямо сражение. Останките са датирани към 1250 – 1200 г. пр. Хр.
От 2008 г. археолозите проучват археологически обект в долината на р. Толензе (десен приток на р. Пеене) в североизточната част на Германия.
В това проучване са участвали и водолази, които са изследвали дъното на речното корито покрай обекта, който е с размери около 2 квадратни км.
Данните за проучването са публикувани в списание „Античност”( http://antiquity.ac.uk/).
Изследователите са открили останките на около 100 човешки тела (предполага се, че жертвите са били над двойно повече!), от които някои имат лезии по костите. Повечето от останките, но не – всички, са на мъже на възраст 20-40 години. Уврежданията по черепите им са били причинени от мощни удари или от стрели, а някои от нараняванията са били фатални.
Сред убитите има и скелети на жени и деца.
Такива наранявания на черепите вероятно са следствие от свиреп двубой лице в лице по време на сражение. Възможно е това сражение да се е провело между враждуващи племена, казват изследователите. Причината за схватката е неизвестна.
В раменната кост или в горната част на ръката един от урелите там на дълбочина ок. 22 см. се е забило острие на стрела. А фрактура на бедрената му кост показва падане от кон. Недалеч от неговите останки са били открити и костите на кон. Много от черепите и костите са поразени от удари с тъпи предмети. Много от нараняванията са били нанесени непосредствено преди жертвите да са умрели. Не са открити следи от опити за лечение на раните им.
Мъртвите тела не са били погребани при с нормална погребална церемония, а вместо това са били изхвърлени в реката. Възможно е някои от бойците да са били обградени и изтикани в реката, където са били избити или са се издавили.
Археолозите открили също останки от два дървени боздугана, единият – с формата на бейзболна бухалка, изработен от ясен, а другият – с формата на чук за крокет, изработен от трънка. Това са оръжия, използвани от обикновени хора. Знатните са се биели с мечове.
Д-р Харалд Любке от германския Център за балтийска и скандинавска археология заявява, че останките доказват, че това е бойно поле, може би – най-древното бойно поле, открито до този момент.
Археолозите не откриват керамика, накити или павирани повърхности, които биха били доказателство за някакви древни погребални обичаи.
Голяма част от костите, изглежда са били отнесени на известно разстояние от реката, въпреки че някои останки изглежда са били изоставени в първоначалното си положение. Изследователите предполагат, че немалка част от труповете е била изхвърлена в реката, която ги е отнесла надалеч, където вероятно са се отложили заедно с пясъчните наноси.
Д-р Любке смята, че основните бойни действия са се провели някъде по горното течение на реката, а откритите останки представляват само една малка част от избитите в сражението. Той се надява да открие още много останки, които биха дали по-точна картина на кръвопролитието.
„Абсолютно необходимо да бъде открито мястото, където труповете са били изхвърлени в реката. Това би дало яснота дали става дума за сражение, за човешко жертвоприношение или за нещо друго. Но ние вярваме, че сражението е най-доброто обяснение за случилото се тук.”, казва археологът.
Доказателства за версията за сражение са както бойните наранявания (пробити или счупени черепи, връх на стрела, забит в кост, строшени кости, счупени прешлени, лезии (следи от остриета) по костите, също така – и наличието сред част от човешките останки на просо – храна, която не е била характерна за Северна Германия по това време, което най-вероятно доказва, че част от избитите са били нашественици.
Тъй като гъстотата на населението по това време е била около 5 души на квадратен км., това би трябвало да е било най-значимото сражение по това време в земите на днешна Германия.
По онова време долината на реката е била заблатена; именно блатният торф е запазил останките от древната трагедия. По онова време растителността на това място е била съставена предимно от треви, тръстики и храсти.
Планира се археологическият обект да бъде изследван до 2015 г. Предстои да се направи ДНК-тест на част от останките.
Археологът се надява археологическият обект да разкрие своята кървава тайна, а неговите открития да дадат тласък на т. нар. археология на бойните полета (Battlefield archaeology).
Към края на 2008 г. отново в Северна Германия, недалеч от градовете Гьотинген, Хановер и Нортхайм археолози открили „находката на века” – бойно поле, при археологическите разкопки на което било открито бойното снаряжение на цял римски легион от III век сл. Хр. (ок. 200-250 г.)
Находките са над 600 метални артефакта, предимно – оръжия, брони, шлемове, стрели, чиято дървена част е от дърво, произхождащо от Северна Африка, метални върхове за стрели със следи от ДНК върху тях, „нокти” от подметка на римски военен сандал, болтове за катапулти и балисти… Открити са и над 300 снаряда (каменни и железни топки) за скорпион (вид катапулт) и копия за балиста. Археолозите веднага се сетили за началната батална сцена във филма „Гладиатор”.
Според историците откриването на това бойно поле е значимо археологическо събитие, понеже то опровергава предположението, че римляните са преустановили опитите си да завладеят Германия след унищожаването на три техни легиона от германите в Тевтобургската гора през 9 г. сл. Хр. Това е интересен щрих от историята на древен Рим.
Откритието било направено от иманяр, който незаконно „изследвал” средновековна крепост в района и, понеже римската епоха не го интересувала, уведомил властите, че е попаднал на нещо, което ще ги впечатли.
Археолозите са убедени, че това са следи, сочещи присъствието на римски военен контингент от поне 1000 легионери. Според тях не е възможно германите да са закупили това оборудване от римляните. Откривани са много находки от въд римския limes (държавната граница), но попадането на бойно поле било голяма изненада за специалистите.
Ариус (280 – 336), либийски презвитер, живял в Александрия, нарушава каноните с твърдението, че Бог Отец е старши в Светата Троица и сам е сътворил Logos (Словото) – автор на нашия свят. Независимо че този възглед среща силна съпротива, Ариус бързо печели и много последователи, сред които двама епископи, седем презвитери, дванайсет дякона и седемстотин монахини. Смутена от силата на тази секта, църквата отнася въпроса към Вселенския събор, свикан през 325 г. в Никея. Там арийците признават, че отхвърлят теорията за единството на Бог Отец и Бог Син, и съответно учението им е обявено за ерес.
Гностици
През втората половина на I век, когато християнството се разпространява и далеч извън пределите на Юдея, някои християни предимно в Сирия и Египет се отказват за пръв път от юдейската част в тази религия. Те осмиват Стария Завет и бога на евреите и възхваляват Христос като земна проява на истинския Бог. Впоследствие се разделят на многобройни школи, в които са заимствани идеи от зороастризма, питагорейството и гръцката философия. Коринт (първата половина на I век) е един от основателите на сектата на гностиците. Тя среща силна съпротива от страна на ортодоксалната църква и след III век постепенно отмира.
Копти
Монахът Евтих (след 440 г.) дръзко изопачава ортодоксалната вяра. Той проповядва, че Исус Христос има само божествена/единствена природа и никога не е добивал телесен/материален облик от девственицата Мария. Сектата е отлъчена от Вселенския общоцърковен съвет, свикан през 451 г., но успява да оцелее в Египет като египетско християнство или копти.
Несторианци
Нестор е монах от Антиохия в Западна Азия, който става патриарх в Константинопол. В края на двайсетте години на V век той започва да проповядва разграничаване на божествеността на Исус от човешката природа на Христос. Това му позволява да отрече възможността девствената Мария да е майка на Бог и младенецът Христос (въпреки божествената природа, която му се приписва) да е достоен за преклонение обект. Тези убеждения вбесяват Кирил, патриарх в Александрия, който организира благочестивата съпротива на всички правоверни християни от Египет и Италия. На Вселенския събор, свикан през 431 г. в Ефес, Кирил успява да постигне обявяването на възгледите на Нестор за ерес, като урежда гласуването да стане преди пристигането на повечето от привържениците на Нестор.
Албигойци
Гностиците изчезват, но след като са дали основа за развитие на друга по-жизнеспособна секта, манихейството, прераснало в албигойската секта (по името на град Алби във Франция), широко разпространена в Европа през XII век.
Англиканска църква
След радикалния теолог Джон Уиклиф (1320 – 1384) много от християните в Англия оспорват върховенството на римокатолическата църква. Независимо от това те формално остават в лоното й чак докато крал Хенри VIII се възкачва на трона (през 1509). Хенри VIII се жени за своята втора жена Анна Болейн през 1533 г. Английската църква обявява предишния му брак с К-терина за недействителен. Но не минава много време и папата се произнася с противното становище – валиден е първият брак. Това изправя краля пред твърде сложен проблем. Решението на Хенри VIII е просто и ефективно – той анулира върховенството на римокатолическата църква за територията на Англия, лишава дъщерята на Катерина от наследство и обявява англиканската църква за самостоятелна организация, оглавявана от него самия. Тези, които не са приели новата структура, са осъдени на смърт. В допълнение новата доктрина е разгласена, а старата вяра – преследвана. Ражда се англиканската църква.
Лютерани
За повечето хора Лутер олицетворява Реформацията в Църквата през XV век. Но през 1529 г. той подхваща остър спор с други реформатори относно значимостта на Тайната вечеря. Неговите последователи в този спор са наречени „лютеранци“ и от този момент може да се идентифицира самостоятелна Лютеранска църква.
Реформаторски църкви
Опонентите на Лутер в спора от 1529 г. основават този клон в протестантството, наречен по-късно Реформаторска църква. Техен лидер е шведският теолог Улрих Звингли (1484 – 1531). Те твърдят, че тялото на Христос всъщност не може да бъде представено посредством хляба и виното по време на Светото причастие, и следователно нямат необходимите общи възгледи с лютеранците, за да останат в една и съща църква.
Протестанти
Повечето от по-късните разклонения на Църквата се обединяват от общото име протестанти. Отначало това е било оправдано, защото всички те са били обединени от протест срещу декрет на Втория международен конгрес, състоял се през 1529 г., прокълнал всяко ново развитие в религията.
Първият протестантски водач е Мартин Лутер (1483 – 1546), който през 1511 г. публикува във Витемберг (в родната му Германия) своето изобличение за папската система за раздаване на индулгенции на грешниците. Лутер отхвърля върховенството на папата и управлението на църковните съвети. Той изтъква Светото писание като единствен авторитет за християните. Лутер отхвърля принципа за безбрачие на християнските свещеници като привнесен по-късно към вярата и сам дава пример, като се жени за калугерка.
Калвинисти
През 1533 г. френският ученик Жан Калвин (или Ковин) (1509 – 1564), преоблечен като търговец на вина, напуска Париж, където възгледите му на протестант са предизвикали гнева на властите. След това той много пътува и пише – резултатът е създаване на калвинизма. Първата калвинистка църква възниква от Реформаторския събор в Женева, чието управление Калвин е поел през 1537 г. Доктрината на калвинизма в действителност съвпада в голямата си част с написаното от Свети Августин (354 – 430), римски теолог от Северна Африка, в „Столицата на Бог“. Допълнението, наложено от Калвин, е изискването за пуританско поведение от всички посветили се на похода срещу греха.
Баптисти
През началото на XVI век една секта, наречена анабаптисти, набира сила в Европа. Тя проповядва, че младенците не трябва да се кръщават, защото малките деца все още не разбират смисъла му. Те смятат всички блага за общи, включително и жените. Анабаптизмът е разгромен – през 1535 г. последователи на протестантите атакуват и разбиват неговата главна квартира в Мюнстер – Германия. През периода 1530 – 1550 г. умерени последователи на същата секта основават една по-приемлива за обществото църква в Холандия. Те запазват своите възражения срещу кръщаването на младенците, но за всичко останало възприемат доктрините на Реформаторските църкви. Основатели са художникът върху стъкло Давид Джорис (1501 – 1556) и свещеникът Мено Симонс (1496 – 1561). Известни са в Холандия като менонити, в Германия – като тауфгесинти, а в англоговорящите страни – като баптисти.
Квакери
Джордж Фокс (1624 – 1691) е един обикновен 19-годишен чирак обущар в селската част на Англия, когато духът му подсказва да изостави шилото и да приеме битието на странстващ мистик. В Манчестър през 1648 г. той се захваща с приспособяване на християнството към собствените си възгледи. Той проповядва присъствието на Христос в сърцето на всеки, като тази вътрешна светлина измества всички външни източници за религиозна вяра, включително Светото писание. Безредията, предизвикани от поведението му, го водят към затвора на Манчестър – първото от много следващи посещения на затворите заради проповедите му. Името „квакери“ последователите му получават през 1650 г., защото Фокс им заповядва да треперят при споменаване на думата Бог. Името, което самите те си дават, е Обществото на приятелите.
Методисти
Сектата на методистите е основана в Англия от Джон Уисли (1703 – 1791), едно от деветнайсетте деца на англиканския пастор Самуел Уисли. Джон Уисли изказва пожеланието „Божието име не трябва да се споменава повече, иначе ще потъне във вечно забвение“. Той няма намерение да основава нова секта, а по-скоро да каже това изречение на хората, чиято вяра е пренебрегвана от регистрираната църква. Този акт има толкова силно въздействие, че Уисли спечелва много последователи – в Британия, Азия, Америка и островите на Тихия океан. Появява се нова църква.
Мормони
Много са забележителните религии, създадени в Съединените щати на Америка, все пак не са много тези, които имат свръхестествено начало като това на „Църквата на Исус Христос и Светците от Късния ден“ -наречена мормони. През 1823 г. един американски ангел, наречен Морони, посещава Йосиф Смит (1805-1844) от Ню Йорк. Ангелът разказва за книгата на Мормон и му разрешава да я изрови през 1827 г. Смит успява да разчете йероглифите с помощта на специални очила, които намира заедно с книгата. За да направи английския превод, той използва камък във формата на яйце, покрито с шоколад. Смит и неговите последователи основават новата църква в Манчестър, Ню Йорк през 1830. Вярата включва някои свойствени догми – една от тях е, че краят на света е много близо. Една от задачите им е да основат нов Йерусалим, но в Америка, и то в Солт Лейк Сити.
Тайпин
Религията Тайпин е създадена в Кантон. Двайсет и три годишният студент Хюн Сюшоан, след като е скъсан за пореден път на изпит, се разболява. В съня си той разбира, че е по-малкият брат на Исус Христос, а мисията му е да унищожи злото и фалшивата религия. През 1843 г., след като е разбрал напълно смисъла на съня си, той основава обществото на „Богомолците“ и започва да проповядва нов вариант на християнството – отхвърлящ иконите, алкохола и наркотиците, величаещ комунизма и равенството между мъжа и жената. Бог е дал на своя по-малък син задачата да опровергае фалшивите вероизповедания на будизма и даоизма и нечестивото светско учение на Конфуций. Привържениците на Тайпин са десетки милиони. През 1850 г. те вдигат въстание в Китай. През 1863 г. Хюн търпи провал в мисията си, след като бунтовниците от Тайпин са разбити от армия, водена от последовател на Конфуций.
Адвентисти от Седмия ден
През 1831 г. американският проповедник Уилям Милер основава движението на адвентистите от Седмия ден. Милер учи последователите си да се въздържат от алкохола и наркотиците, да направят почивен ден събота и че краят на света ще настъпи през 1843 г. Движението приема неговото име през 1860 г., а се конституира като църква през 1863 г.
Свидетели на Йехова
Чарлс Ръсел основава сектата във Филаделфия през 1872 г. Тя съчетава проеврейска активност с вярата, че Исус е бил обикновен смъртен, съвършен човек, но не и божествен. Неговите „Международни ученици на Библията“ не празнуват рождени дни, включително Коледа, нито признават зависимост от светската/временна власт. Те приемат името „Свидетели на Йехова“ през 1931 г.
Обединена световна църква
През 1936 г. Исус Христос се явява на шестнайсетгодишния Сун Мюн Мун от Северозападна Корея и го кани да завърши делото Му на Земята. Обстоятелствата пречат на младият Мун, докато е в Северна Корея. Той бяга на юг през 1950 г., където успява да формира малка група от последователи. През 1954 г. създава „Асоциация на Светия Дух“, сега широкоизвестна като „Обединена световна църква“ или „Мунисти“/“Църквата на Мун“. Нейната задача е да избистри идеите в световното християнство и да обедини християните от целия свят зад веруюто на Мун и под неговото ръководство. В Корея, Япония и Америка тази секта има много последователи и членове.
Отначало металните продукти са се правели от самородно сребро и злато, но се използвала само груба техника, като изчукване и издълбаване. Металургични техники са въведени за пръв път при производството на олово.
Топене
Оловото не се среща в естествена метална форма, но богатата на олово руда, галенитът, има такъв метален вид, че праисторическите експериментатори са се изкушавали да отделят метала от нея. Това те са направили за пръв път през VII хилядолетие пр. Хр., като са открили топенето. Подлагайки галенита на много високи температури, постигани в керамични пещи, те са добивали разтопено олово. Малки прости оловни предмети, датиращи от онова време, са намерени в Чатал Хуюк, процъфтяващ праисторически град в днешна Турция, а също и на места в Северна Месопотамия (днешен Ирак). Една от тези страни е първата родина на топенето отпреди повече от 8000 години.
Леене
Топенето вероятно е проправило път на друга фундаментална техника, леенето. Тъй като разтопеният метал при охлаждане се втвърдява и приема формата на съда, в който е излят, металурзите започнали да правят отливки с намерението металът да заеме желаната от тях форма. Това е направено за пръв път вероятно в Западна Азия около 4500 г. пр. Хр. или един или два века преди това. Но едва до 4000 г. пр. Хр. стопилките са се отливали само в най-прости форми.
Мед
Тъй като медта се среща в природата в доста чиста метална форма, хората започнали да я обработват, преди да е започнало топенето. Откритите малки медни предмети са отпреди 6000 г. пр. Хр. В Чатал Хуюк са изкопани медни обекти от този период, включително тежести за дрехи. Медта трябва да е влязла в употреба, след като хората забелязали, че изковаването или лъскането на тъмните парчета им придава ярък металически вид. След това те открили, че медта е достатъчно ковка, за да се обработва студена, и започнали да правят малки инструменти, тежести и гвоздеи. Като се има предвид както технологичното ниво, така и наличието на естествена мед, Западна Азия вероятно е родината на медодобива.
Бронз
Бронзът е сплав от мед, която първоначално била направена с добавка на калай. Бронзът с достатъчно калай се обработва значително по-добре и се използва за повече цели отколкото медта. Обаче първите производители на бронз – в Месопотамия и Мала Азия (днешен Ирак и Турция) – явно не са били наясно с това, защото са добавяли съвсем малко калай, така че техният бронз има минимални преимущества пред медта. Предмети, изработени от този долнокачествен бронз (често една част калай на петдесет части мед), са открити в руините на Ур и други месопотамски градове от времето на шумерите, около 3500 пр. Хр. Но липсата на подходящи калаени руди по тези места може да означава, че най-старият бронз в действителност бил направен по-далеч на север. Били са необходими около хиляда години за получаването на истинския бронз (около една част калай на десет части мед), за да се постави началото на „бронзовата епоха“.
Желязо
Ограничено количество метално желязо се среща в естествен вид в метеоритите. Първите железни предмети са направен от такова желязо през IV хилядолетие пр. Хр. Най-старите предмети от разтопено желязо, открити от археолози, са от Месопотамия и Анадола (в днешен Ирак и Турция) и са произведени около 3000 г. пр. Хр. или малко след това. Според местоположението на тези древни находки и разпределението на рудните залежи вероятно топенето на желязото е започнало малко преди началото на IV хилядолетие пр. Хр. най-напред в Анадола, западно от Таврийските планини, в днешна Турция или Армения.
Стомана
От 1400 г. пр. Хр. тази прародина на желязото (вж. по-горе) била под управлението на империята на хетите. Със своите превъзходни руди тя била център за експериментиране в металургията на желязото, развивайки техники, твърде различни от използваните при бронза. Това експериментиране довело до производс-твото на първата стомана, заслугата за което е на един народ от Северен Анадол, наречен чалиби. В продължение на около два века превъзходните руди и технологията дали на империята на хетите практически монопол в производството на желязо и стомана и експортната търговия с желязо.
Висока пещ
Но тази стомана не стигала до Китай и китайците трябвало сами да изобретят топенето на желязото, което станало доста бавно. В Китай няма следи от желязо, датиращо отпреди около 600 г. пр. Хр. И въпреки това две столетия по-късно китайците вече са въвели високата пещ – една стъпка, която Западът направил двайсет и седем века по-късно. Преди това изобретение желязото трябва да е било произвеждано във формата на ковано желязо, а стоманата била добивана чрез увеличаването на съдържанието на въглерод.
Висока пещ
Лято желязо
Високата пещ позволила на китайците да добиват лято желязо, което има високо начално съдържание на въглерод. Най-старото открито засега желязо, отлято от висока пещ, представлява блок от 500 г. пр. Хр., който е намерен близо до Китай. Тайната на китайския успех може да е била в това, че те са имали руди, богати на фосфор, или са прибавяли фосфор при зареждането на високата пещ. Постепенно проектирането и операциите по обслужването на високите пещи се подобрявали и технологията се разпространила в Индия и Япония.
Лята стомана
Китайците произвеждали стомана чрез обезвъгляване на лято желязо. За да направят процеса по-ефективен, през II в. пр. Хр. те започнали да продухват горещото още разтопено желязо с окисляваща струя студен въздух. В Китай и Япония през XVII век (най-късно) това довело до процес за производство на лята стомана, който включвал подлагане на разтопената маса от желязо на действието на студена въздушна струя. (Това е първата продукция на лята стомана по метод, който засега е неизяснен. Съвременните китайски археолози съобщават обаче, че стомана се е произвеждала още по времето на първата династия на Хан – 206 г. пр. Хр. – 9 г. след Хр. – чрез топлинна обработка за обезвъгляване на твърдо лято желязо. Това обаче се нуждае от някакво обяснение, преди да го приемем.)
Неръждаемо желязо
В Делхи има колона от желязо, която стои на открито от около 400 г. след Хр. и не показва следи от ръжда или разрушение. Тя се състои от 99,72 % желязо, 0,08 % въглерод, 0,046 % силиций, 0,006 % сяра и 0,114 % фосфор. С възможностите си да произвежда желязо с такъв състав индийската промишленост изпреварва Европа с петнайсет века, а и самата формула е разкрита само чрез анализ на колоната. Причината за нейната издръжливост е спорна. Едно от предположенията е, че това се дължи на ниското съдържание на сяра и липсата на манган. Техниката не може да е много по-стара от колоната, защото иначе и по-ранни образци биха издържали до днес. Но колкото и да е поразително това желязо, то едва ли ще продължи да се произвежда след изобретяването на неръждаемата стомана.
Неръждаема стомана
Неръждаема стомана се нарича сплав от желязо и хром, която издържа на корозия. Първата такава стомана е произведена експериментално през 1904 г. от френския учен Леон Гиле. Любопитно е, че Гиле не е съобщил за забележителната устойчивост на корозия на своята нова сплав и че неговото откритие не е било използвано веднага. От 1912 до 1914 г. Едвард Маурер и Бено Щраус от германските заводи за производство на стомана „Круп“ в Есен разработили астенитна неръждаема стомана с голямо съдържание на никел. Свойствата й са идеални за нуждите на индустрията и така тръгнало нейното производство в големи количества.
Месинг
Месингът е сплав, която се състои главно от цинк и мед. Древните гърци и жителите на Западна Азия преди почти 3000 години са получавали месинг чрез стопяване на руда с правилно съотношение на двата метала, без да са разбирали неговия състав. Първият месинг, получен при изкуствено смесване, идва от Мала Азия (днешна Турция) или от Егейските острови, около 400 г. пр. Хр. Теофраст (около 383 – 287 г. пр. Хр.), ръководител на Лицея в Атина, в книгата си „De Lapidibus“ споменава, че за тази цел е използвана цинковата руда каламин. (По същото време месингът е получен независимо и в Индия, но индийският не е датиран достатъчно точно, за да се знае дали предшества западния.)
Цинк
Гръцка книга по география, публикувана в Римската империя през I в. пр. Хр., споменава за метал, наречен псевдосребро, което трябва да е цинк, понеже се казва, че е съставка на месинга. Това би означавало, че цинкът е получен в чист вид някъде на запад – може би в Египет или Западна Азия, вероятно около 100 г. пр. Хр. или по-рано. След това методът за производството му е загубен до 1797 г., като изключим Индия и Китай, където той е изобретен отделно.
Мелхиор
Мелхиорът, известен преди това като пакфонг, е сплав от мед и никел, от която обикновено се правят монети. Нейното използване за пръв път е доказано в монетите от елинистичната епоха на Бактрия (в северната част на днешен Афганистан, на сухоземния търговски път между Китай и Персия) около 200 г. пр. Хр. Източникът на метала е все още неизвестен, но вероятно е внесен от Китай. Монетите на Бактрия имали една част никел и пет части мед, подобно на стария китайски мелхиор, който обаче китайците са започнали да използват за производство на монети едва през VI в.
Никел
Подобно на цинка никелът започнал да се произвежда като чист метал дълго след като е бил използван като сплав. Шведският химик Аксел Кронстед (1722 – 1765) пръв получил никел през 1751 г., като го извлякъл от сулфиди, отделени от кобалтова руда за целите на производството на кобалтово-синя боя.
Алуминий
За наличие на метал в алуминиевата руда се е подозирало много преди да е известно някой да е успял да го извлече. През 1825 г. откривателят на електромагнетизма, датският физик Ханс Кристиян Оерщед (1777 -1851 ) доказал, че подозрението е вярно. Оерщед не разработил надежден метод за извличане. През 1846 г. германският химик Фридрих Вьолер открил, че чрез нагряване на смес от натрий и алуминиев хлорид може да се извлече чист метал във вид на многочислени миниатюрни капчици.
Около 450 г. пр. Хр. Левкип от гръцкия град Елея в Южна Италия стига до извода, че Вселената е съставена от миниатюрни невидими частици, които той нарича атоми. Съгласно неговата книга „Великият световен порядък“ Вселената е неограничена по размери, но се състои от празно пространство, частично запълнено от постоянно движещи се атоми. Според него тези атоми са елементите, които, когато се групират по различен начин помежду си, образуват цялото многообразие на наблюдаваните вещества.
Около 450 г. пр. Хр. Левкип от гръцкия град Елея в Южна Италия, който стига до извода за атома има последовател – това е Демокрит от Абдера, Северна Гърция. Народът на Абдера бил нарочен за глупав, но Демокрит е един от най-умните мъже на всички времена. Той е наричан „смеещият се философ“, защото проповядвал, че човек трябва непрекъснато да е в добро настроение. Неговият основен принос към науката е в откриването на атомната теория и обяснението на природата на химичните съединения. Демокрит твърди, че разликата в качествата на материалите (топлина, вкус, твърдост) не се дължи на единичните атоми, а на тяхната подредба и степен на възбуда, и особено на начина, по който са свързани. Формата и външността на различните видове атоми им позволява да се свързват по множество различни начини, като така се образуват материалните вещества. Той обозначава различните атоми с букви от азбуката, които образуват думи, и по този начин илюстрира идеята за молекулата. Независимо че Демокрит допуска разрушаване на междуатомните връзки посредством външна сила, той не допуска разцепването на самите атоми.
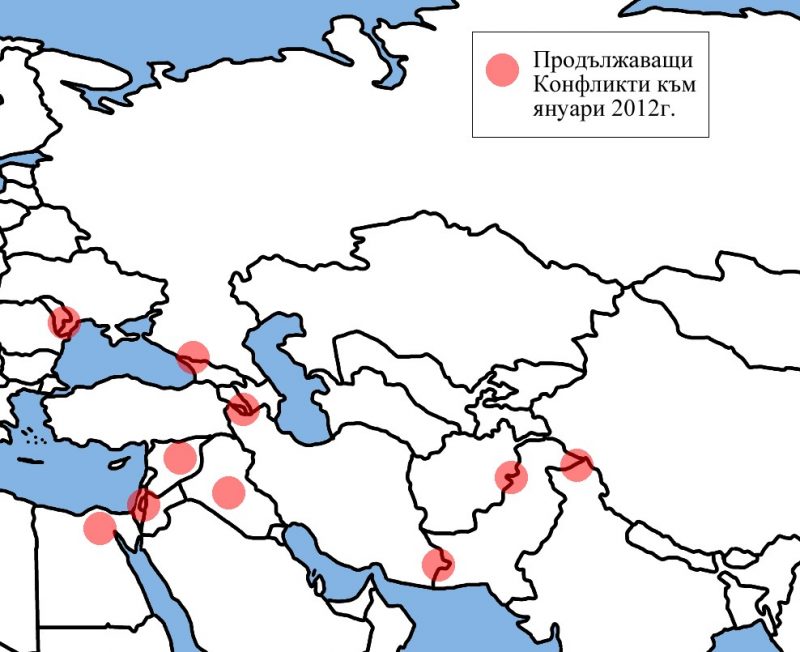
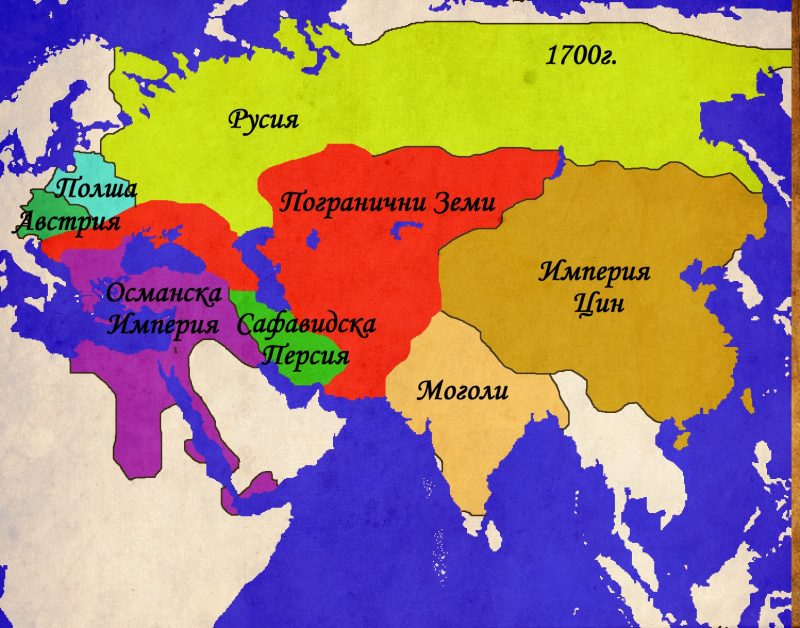
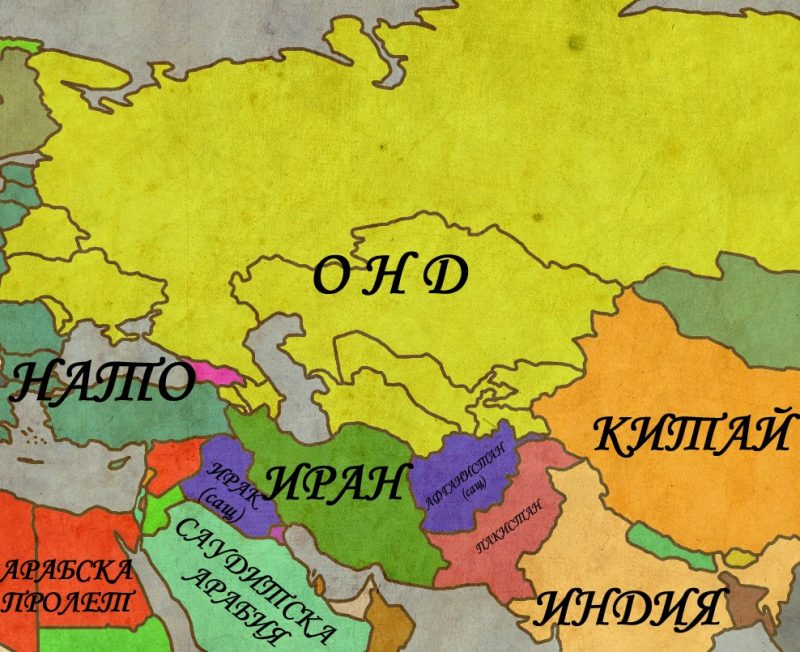
Централна Евразия в политиката на Великите (1500-2012г.)
Последните пет столетия са свидетел на постоянните опити на различни империи от всички краища на света да наложат ако не директната си власт, то поне влиянието си върху колкото се може повече територии. Има, обаче, един обширен район на света, където много от големите империи са изпробвали силите си, проваляли се е и този провал повече или по-малко е коствал съществуването им. Този район може най-общо да бъде наречен Централна Евразия и настоящата статия има за цел да направи един обобщен преглед на опитите на Големите да доминират над тази част от света.
Тъй като това не е обикновена статия, предназначена за публикуване в блог или в пресата, е нужно да започнем по-отдалеч и да изясним термините с които ще се борави в текста по-долу. Ето защо, вместо увод, следващите редове ще отговорят на няколко основни въпроса, без които читателите (а и автора) не биха могли да продължат по-нататък към същинската част.
Какво е Централна Евразия?
С термина Централна Евразия, в настоящата статия ще се разбират териториите разположени между Черно море на запад и Тибет на изток. Те обхващат разнообразни по климат, предимно планински райони, населявани още от дълбока древност от свободни, независими племена, кланове и родове, които трудно понасят съществуването на централизирана държава и се стремят да запазят максимална възможна автономия спрямо властовите центрове . Защо Евразия? Тук искам ясно да отбележа, че терминологията няма нищо общо с евразийството, което е популярно идеологическо течение в Русия, което се стреми да фиксира мястото на тази страна като мост между културите на двата континента. Настоящата статия възприема идеята, че Европа и Азия образуват един по-голям комплекс не само в географско, но и в културно и политическо отношение. Излишно е да се навлиза в детайли относно това как големите културни и политически процеси започнали в единия край на Евразия засягат целия комплекс. Два христоматийни примера са формирането на Монголската империя и Великото преселение на народите, но има и безброй други, които също могат да бъдат посочени. Важното в случая е, че Европа и Азия са до такава степен свързани по между си, че изкуственото географско деление, наложено от европейците има малко общо с действителността. Приемайки Евразия като факт, настоящата статия ще се фокусира върху нейната централна част, която през по-голямата част от новата и най-новата история се оказва в периферията на големите империи.
Какво е империя?
Това е може би най-трудния въпрос в съвременната историография. Изписани са множество статии, сборници и монографии и доста от тях предлагат често противоречиви дефиниции и параметри по които можем да отсеем империите от останалите политически формирования. Въпросите за размера, етническото многообразие, експанзивността и идеологията са също толкова важни колкото и самоопределянето на една държава като империя, както и начина по който съседите й я възприемат. Отделно от това съществуват множество религиозни и лексикални особености, които няма как да бъдат прескочени. Лично аз съм противник на идеята, че властови и религиозни термини, които не са продукт на западно-евразийските култури трябва да бъдат превеждани и приравнявани към стандартите на тези култури. Веднага давам пример с опита на британците да унифицират китайските дворцови титли със своите или опита за приравняване на титлите „халиф“ и „император“. Всеки средно интелигентен човек знае, че два народа могат да влагат коренно различно значение в на пръв поглед идентични термини, позиции или явления. Ето защо е трудно да се наложи единен стандарт за това кои държави са империи и кои не са. За да избегнем дълбоките и противоречиви дебри на научния спор, тази статия ще използва един по-свободен модел за „империя“. Под империя от тук на сетне ще се разбират големите колониални държави, заместени през ХХ-ти век от т.нар. Велики Сили. За още по-голямо улеснение ще изброим конкретните държави – Великобритания, Русия (След 1917г. СССР, след 1991г. Руска федерация), Китай (империите Минг и Цин, република Китай и Китайската народно-демократична република), Османската империя (след 1923г. – Турция), Персия (След 1979г. Иран), САЩ , Моголската империя (чието място днес се заема от Индия). Това, разбира се, не са единствените империи, но останалите подобни държави като Германия и Франция нямат директно отношение към разглеждания проблем и няма да са предмет на настоящата статия. Именно изброените по-горе държави ще са главните герои на нашата история и чрез развитието на тяхната политика спрямо Централна Евразия ще проследим как точно опитите за контролиране на региона влияят на съдбата на империите.
Какво е „властови център“?
Без да претендирам за оригиналност или някаква форма на свръх иновация, смея да заявя, че термина властови център не е повлиян от конкретно научно изследване, а е плод на поредица от лични размишления върху проблемите на гео-политиката. Той е изключително подходящ за да предаде реалното състояние на политическата действителност в Централна Евразия. Какво имам предвид? Когато говорим за обширните територии между Черно море и Тибет, където географските фактори правят комуникациите и директния контрол трудно осъществими, се наблюдава тенденция за формиране на властови центрове, които се опитват да наложат контрол над нестабилната периферия около тях. Пример за това са поредицата от персийски династични империи, Моголската империя, както и държавите на грузинци, афгани и узбеки. Характерно за властовия център е консолидирането на властта около един икономически и политически център, който се превръща в столица на формиращата се държава. В следствие на богатствата и ресурсите осигурени чрез този център, власт-имащите се заемат да разширят влиянието си за сметка на периферните райони, които отстъпват по богатство, брой на населението и наличие на организирани военни сили и административен апарат. Способността на властовия център да задържи определено пространство под своя директен и индиректен контрол преминава през периоди на възход и падение, които често съвпадат с династични промени или изчезване на едни държави и замяната им от други. От другата страна на властовия център имаме периферията, съставена от разпокъсани автономни владения/племена, които се опитват да се противопоставят на домогванията на един или повече властови центрове. Тези племена/владения имат склонност да се превръщат във военно-политически ренегати, да се съюзяват със силите на деня в опит да извлекат максимална изгода или просто да запазят съществуването си. Когато на лице е един географски фрагментиран регион, логично е да се формират множество властови центрове и периферии, чийто мащаб варира в зависимост от човешките и материални възможности на замесените страни. Освен това, отделни властови центове могат да се окажат в ролята на периферия на по-големите от тях властови центрове. Този сложен йерархичен комплекс превръща политическата ситуация в Централна Евразия в изключително проблематична и хаотична материя, чието правилно разбиране изисква полеви опит, сериозно наблюдение и детайлно познаване на местните традиции, вражди и обичаи. За да направи ситуацията още по-заплетена, освен комплекса от съподчинени властови центрове и периферии, в Централна Евразия се наблюдава още един феномен, който е подчинен на свой собствени закони – пограничните земи (The Frontier).
Какво е „погранични земи“?
Терминът „Frontier” използван в англоезичната литература за да обозначи територията, която се намира на ръба на цивилизациите е изключително комплексен. Буквалният превод „граница“ е неподходящ, тъй като предполага една строго фиксирана зона от няколко километра, която разделя две държави. Погранични земи е далеч по-подходящ превод, тъй като предполага един по-обширен географски и политически ареал. Пограничните земи са феномен който обикновено се наблюдава в периферията на сбор от държавни формирования, на по-големи държавни образования или на границата между две империи. Типичен пример за подобни зони са Мала Азия и Кавказ, които се формират като погранични зони през Средновековието в контекста на Арабо-Византийските конфликти. Характерно за пограничните земи (или зони) е сливането на културните и политически традиции на двете противостоящи си държави/цивилизации. В горепосочените примери става въпрос за създаване на хибридни ислямо-християнски вярвания, както и на заемки и от византийската и от арабската традиции в политиката и примесването им с местни практики. Така се формира един комплекс от традиции, практики и идеи, които не могат да бъдат отделени едни от други и често са характеризирани като неортодоксални. За хората, живеещи в тези територии, е характерно чувството за независимост, както и формирането на специфичен начин на поведение, базиран на идеите за чест, справедливост и кръвно братство. Пограничните земи са люлка на особен вид племена, чиято връзка се базира на общия интерес и начин на живот и има малко общо с реално родство. Типичен пример за това са казаците, които са продукт на нуждата от създаване на дружини от верни, целеустремени мъже, за които клетвата и побратимяването означават много повече от кръвното родство. Според най-новите теории, Османците се появяват именно като такава група, обединена от харизмата на Ертогрул и Осман , а не от някакъв мним племенен произход. Тъй като Централна Евразия лежи в периферията на множество империи, голяма част от нея представлява една голяма погранична зона, където традициите на християнството, исляма в двете му основни форми, будизма и шаманизма се примесват в една уникална спойка, която формира характерен пограничен етос . Що се отнася до отношението между програничните общества и държавите, на преден план изпъква стремежа на пограничниците да се откъснат от контрола на заобикалящите ги държави и империи. Това води до една своеобразна преходност на „верността“ на дадена погранична група към една или повече държави. Не е чудно тогава, че в два последователни конфликта едно и също погранично племе (или група) може да воюва на срещуположни страни за да запази собствените си интереси. Тази флуидност на верността прави окончателното покоряване на тези групи доста сложно, тъй като прекалената агресия рискува да предизвика пренасочването на пограничниците от влиянието на една сила към друга. Така например Русия, Османската империя и Персия са ту врагове, ту съюзници на народите в Кавказ, в зависимост от това коя от останалите сили се стреми да наложи властта си над целия регион.
Историята на кратко
Комбинацията между мащабни погранични зони и множество имперски властови центрове предопределя постоянното наличие на конфликти в Централна Евразия. Да се разкаже историята на Централна Евразия е непосилна задача за който и да е отделен изследовател. Ето защо следващите редове предлагат кратък обзор на основните събития, без да претендират за пълна изчерпателност. Историята на отделните империи и техните наследници е обект на множество монографии, немалко от които са достъпни за българският читател. По-новите изследвания на англоезичната историография се фокусират по-скоро върху отделни социални политически явления, тяхното разбиране и интерпретиране, докато по-старите трудове както на „западните“ така и на съветските историци са насочени към изчерпателен разказ на събитията за сметка на по-малко анализи. Оптималният вариант за всеки изследовател е да използва и двата типа изследвания за да получи по-пълна представа за хода и същността на събитията.
Сблъсъкът на Империите 1500 – 1923г.
Годината 1500 е един своеобразен вододел в историята на Централна Евразия. Въпреки че тази година не носи особено значение за „западната“ цивилизация, за народите, населяващи земите между Черно Море и Тибет тя се оказва ключова. За какво става дума?
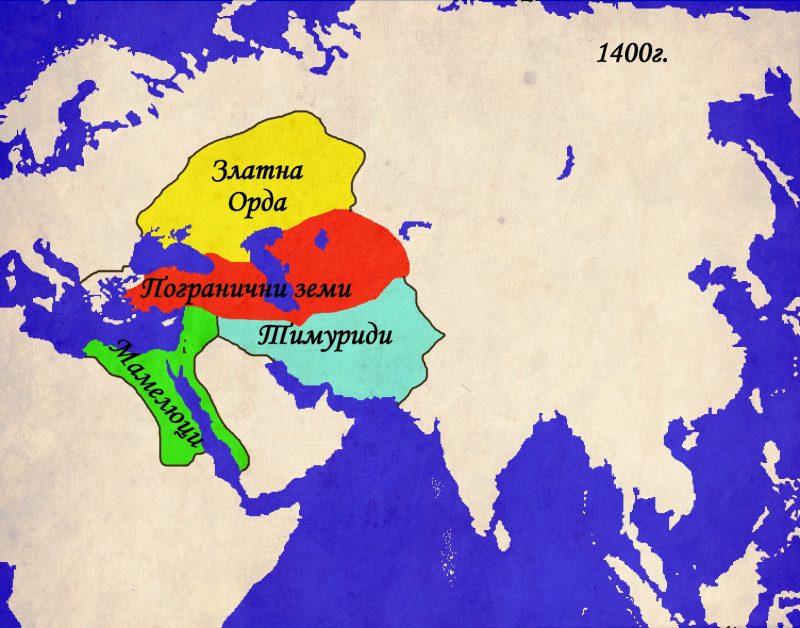
В началото на шестнадесетото столетие, територията на Централна Евразия представлява една сложна плетеница от по-големи и по-малки държави, както и обширни територии, населявани от множество различни племена. Старите хегемони на отминаващото Средновековие са предизвикани от издигащите се сили на Новото Време. Халифатът на Мамелюците в Египет и Сирия все повече се приближава към своя край, изправен пред нарастващата мощ на османците. Последната голяма тюркска държава – Аккоюнлу (Федерация на Белите Овни) губи борбата с полунезависимите племенни вождове, които отказват да следват повелите на султаните на Аккоюнлу. По на изток, последните остатъци от някога могъщата Тимуридска империя попадат под ударите на узбеките. В тази нестабилна обстановка изгрява звездата на двама мъже, които в последствие ще станат близки приятели и съюзници. Единият е последния пряк потомък на Тамерлан, а по майчина линия и наследник на Чингиз Хан. Другият е последният потомък на имамски шиитски род, който е династично свързан с повечето властващи фамилии в региона. И двамата имат тежко детство, изпълнено със заговори, бягства и несигурност. И двамата ще вдигнат падналите знамена на своите наследства и ще им придадат нов блясък и значение. Първият се нарича Захир уд дин Мухаммад Бабур, родоначалник на Моголската династия. Другият – Абу Мустафар бин Сайдар ас Сафави ще остане в историята под името Исмаил I и ще се превърне в първия персиец, управлявал Персия от времето на Ахеменидите.
Съдбата им е отредила различни пътища. След като узбеките, водени от султан Мухаммад Шайбанипревземат Самарканд през 1500г., младия Бабур отказва да води безсмислена война за старото тимуридско наследство и решава да потърси съдбата си другаде. Съдбата изглежда си знае работата и няколко години след като Бабур укрепва властта си над южен Афганистан, тигърът открива своята плячка. Делхийският султанат , управляван през последните век и половина от династията Лоди запада, разкъсван от междуособици. Бабур събира малка, но опитна армия, въоръжена с аркебузи и оръдия и нахлува в Индия. Осланяйки се на по-многобройните си сили, Сикандер Шах Лоди пресреща силите на Бабур. Последвалата битка при Панирпат (1526г.) се превръща в едно от най-значимите събития не само в историята на Индия, но и на Евразия като цяло. Победата на Бабур е категорична, династията Лоди е ликвидирана, а на нейно място се въздига нова империя, която ще властва над Индия, Афганистан и Пакистан през следващите три столетия.
Исмаил използва дезинтеграцията на Аккоюнлу за да консолидира своята собствена власт в южен Азербайджан и Северозападен Иран. Залагайки на мобилността и религиозния фанатизъм на своите тюркмени-казълбаши („червени шапки“), Исмаил започва една война, която ще трае до края на живота му. Отслабените сили на Аккоюнлу не представляват особена пречка за армията на Сафавидите . Истинският опонент се оказват османските армии, водени от своя султан Селим I. Османците вече са изградили най-голямата и опитна армия в Евразия, която, към онзи момент, няма еквивалент по бойните полета. Въоръжени с огнестрелни оръжия и оръдия, османците лесно се справят с предимно кавалерийската армия на персите. Религиозната ярост на еничарите все още не отстъпва на тази на казълбашите, но тюркмените са надиграни и като дисциплина и като въоръжение от „слугите на Портата“. Битката при Халдиран (1514г.) и последвалите няколко десетилетия войни завършват в полза на османците. Персите губят Месопотамия и Азербайджан, изоставяйки столицата си в Табриз за по-отдалечения и защитен Исфахан. Въпреки тези неуспехи, Исмаил успява да нанесе поредица поражения на узбеките както и на владетелите в Източна Персия (Херат, Хвазрим и Бухара) и Балуджистан. По този начин Сафавидите компенсират загубеното на запад с експанзия на изток. До смъртта си Исмаил и Бабур остават съюзници, но постепенно империите им се противопоставят както на религиозна , така и на политическа основа .
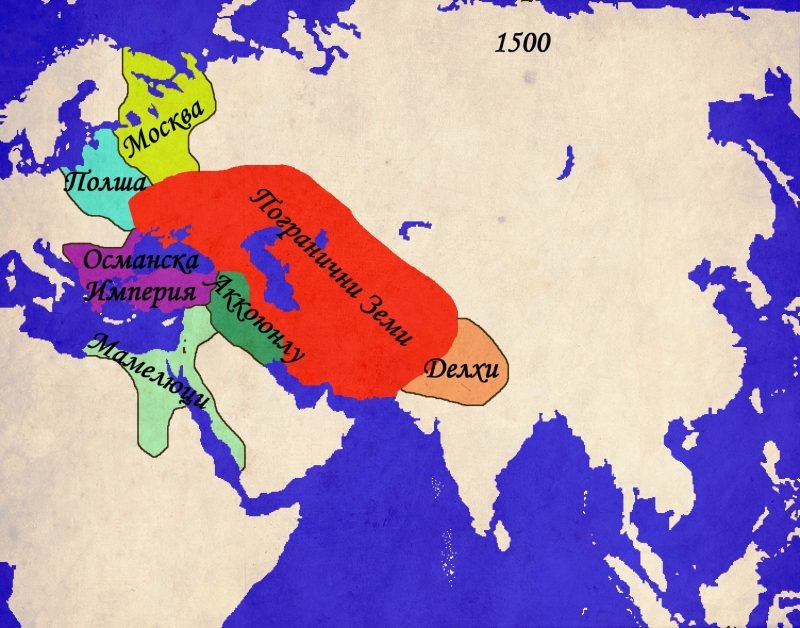
Междувременно западните предели на Централна Евразия постепенно попадат под властта на третата голяма ислямска династия на епохата – Османците. След победите над Сафавидите (1514) и Мамелюците (1517), Османската империя подчинява Месопотамия, Сирия, Палестина, Египет и Хеджаз. Следващи експедиции при управлението на Сюлейман I Кануни поставят Йемен и Еритрея под османска власт. По същото време борбата с Персия продължава и след поредица от конфликти османците фиксират границата далеч на изток в пределите на днешен Иран. Едва след 1600г., в следствие на икономическите и военни проблеми на османците, Сафавидите, в лицето на Аббас I, успяват да отвърнат на удара, завладявайки части от Месопотамия и Азербайджан. След 1630г. Османците поемат инициативата и според мира от 1638г. Сафавиди и Османци установяват граница, която в общи линии ще остане непроменена до 1920г. По същия начин се развива и конфликта между Сафавиди и Моголи. Борбата е за властта над Кандахар, който контролира основния път между Индия и Персия. Града сменя собственика си много пъти между 1600 и 1700г., но като цяло остава в ръцете на Моголите.
На северозапад от разглежданата по-горе зона се появява нов регион, който се превръща в цел на редица имперски апетити. Казвказ и северното Черноморие стават арена на поредица от конфликти между трите големи сили на западната степ – Османците (заедно с техните татарски васали), Посполитажеч и Московското царство. Трите държави и техните локални съюзници се вплитат в поредица от конфликти, които с редки прекъсвания продължават чак до 1783г., когато Русия анексира Кримското ханство и слага край на османското присъствие в северното Черноморие.
По подобен начин се развива и конфликта в Кавказ. Там до 1721г. Основните опоненти са Османската и Сафавидските империи, които си оспорват върховенството над местните държавици и племена. След 1721г. Русия също се включва в надпреварата за Кавказ. Процеса на присъединяване и усвояване на Кавказ се оказва изключително бавен и труден за Русия и остава неразрешен въпрос в руската външна политика и до ден днешен.
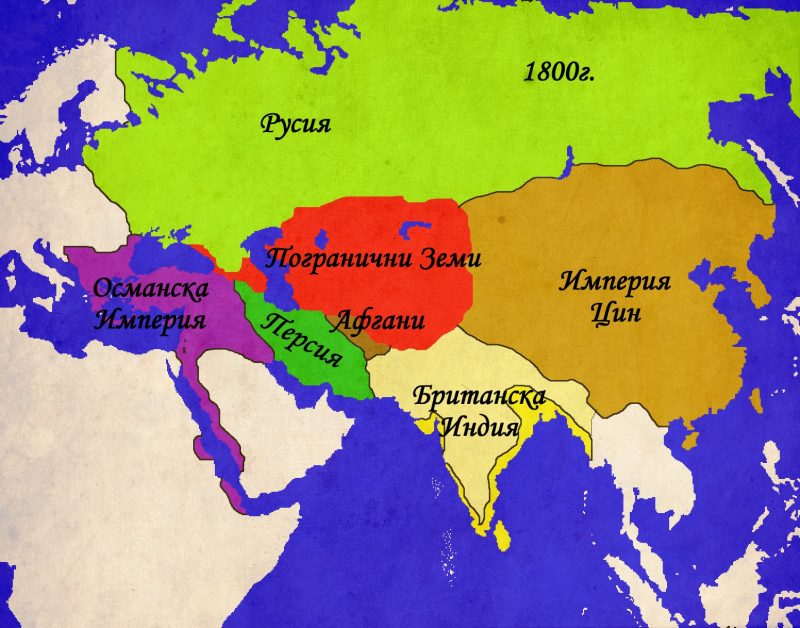
Един последен и по-малко популярен фронт между империите и пограничните земи се намира на териториите източно от Аралско море и северно от Тибет. Тези земи, част от „Пътя на коприната“, от векове са спорна зона между степните вождове, монголските ханове и владетелите на Китай. След краха на династията Юан през 1368г., новата китайска династия Минг се заема да се разправи с монголските си предшественици. Поредица от експедиции ликвидират последните гнезда на юанска съпротива и поемат по стария караванен път северно от пустинята Гоби. Тук китайците се сблъскват с узбеките, чагатаите и ойратите, които парират опитите на Пекин да разпростре властта си на запад. Паралелно с това ресурсите на Китай се хабят в безплодна война с Виетнам, скъпо-струващите експедиции на Зенг Хе, както и за грандиозните строителни проекти на император Йонгле , Експанзията на запад е окончателно изоставена, след като в средата на XV в, степните племена нанасят поредица от поражения на китайските армии и дори пленяват един от императорите. Последвалия период на затвореност на Китай продължава чак до края на XVII в., когато новата манчуйска династия Ци отстранява империята Минг. За разлика от своите предпазливи и затворени предшественици, императорите от Ман Чу се заемат с активна и агресивна политика на запад. Тибет е подчинен, заедно с днешна Монголия,. Степните племена на ойратите са сломени, а калмиките са принудени да се изселят на запад, към земите на долна Волга, където се превръщат в несигурни съюзници на Русия. Мощта на Ци остава неоспорима до средата на XIX в., когато поредица от войни с Великобритания, Франция, Русия и Япония разкриват слабостта на империята. Въпреки това, Китай продължава да владее Монголия и Тибет до 1912г., когато империята е заменена от република и двете територии се отделят като самостоятелни държави.
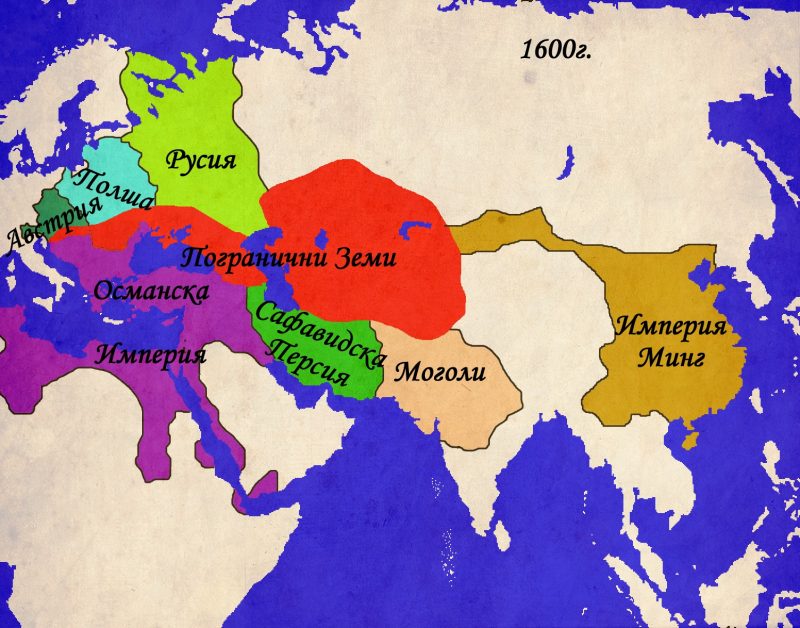
Осемнадесетото столетие носи значителни промени в отношението на империите към пограничните области и по-малките властови центрове. Докато Османската и Персийската империи постепенно залязват, Русия започва да се издига като най-силната държава в Централна Евразия. Използвайки своята по-модерна армия, както и добре развитата система от погранични укрепления, известна като „засечни черти“ , Русия започва да навлиза все по-дълбоко в степите на Евразия. След като подчинява калмиките в началото на века, Русия се заема с подчиня ването на татарите от Крим, както и на техните братовчеди – ногаите. До края на XVIII в. Русия си е осигурила властта над земите северно от Кавказ и западно от река Урал. Войните с Наполеонова Франция забавят процеса на усвояване на степта, но след 1815г., той започва с нова сила. Въпреки това, решителният пробив в Централна Евразия е направен след неуспешната Кримска война(1853-1856г.), когато степта остава единствената достижима цел за руската външна политика. Александър II насърчава експанзията и до 1870г. Русия подчинява всички земи северно от Иран и Афганистан. Паралелно върви и експанзията в Кавказ, където Грузия сама се присъединява към православната империя, последвана от завзетите след 1825г. Армения и Азербайджан. Съпротивата на персите е слаба, но османските сили в Трабзон и Ерзерум оказват постоянна съпротива. Едва след Руско-Турската война от 1877-78г. Русия постига пробив в Кавказ за сметка на отстъпките, направени от териториите на балканските държави, сред които и България.
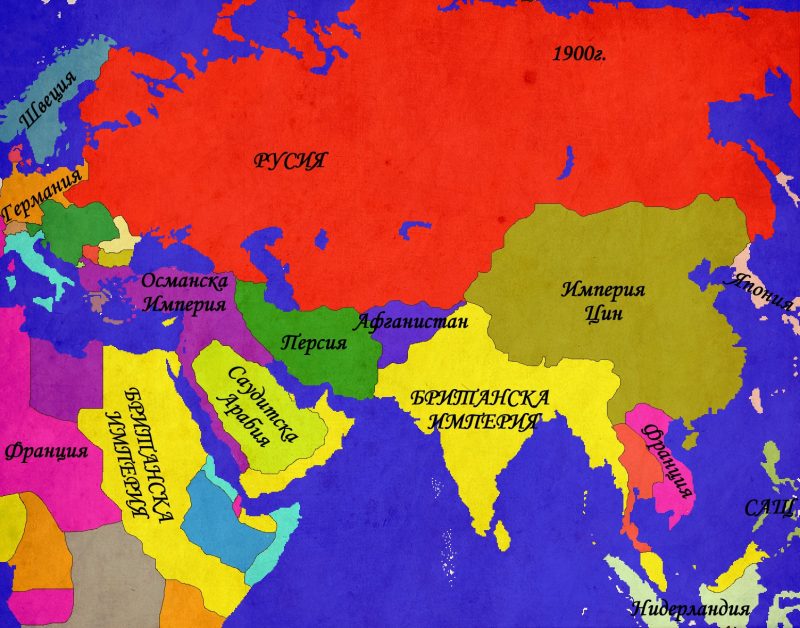
Паралелно с руското проникване, в Централна Евразия се появява още една колониална империя – Великобритания. До края на XVIII в. Британското присъствие се отличава с доминиране на морските пътища, но след Наполеоновите войни, експанзията по суша също се активизира. Нарасналата британска сухопътна мощ, съчетана с все по-бързо развиващата се военна техника позволяват на англичаните, използвайки сравнително малки по численост сили да се справят с големите армии на индийските владетели. Паралелно с това, британците рекрутират все повече местни войници, наричани „сепои“, които обучават по „европейки“ образец и използват в своите войни не само в Индия, но и в останалите части на света. Британската власт достига своя зенит е през 1858г. Когато Източно-индийската търговска компания и нейните владения в Индия са обявени за неотменна част от британската корона. За управлението на Британска Индия (която включва днешните Индия, Бангладеш, Мианмар и Пакистан) е назначен вицекрал, чийто мандат се регулира от парламента и кралицата. След като завзема Пакистан, Британия влиза в пряк конфликт на интереси с Русия, наречен „Голямата Игра“. Двете страни превръщат Афганистан в буферна зона на своите интереси, особено след Кримската война. Опитите на британците да подчинят и Афганистан като превенция срещу по-нататъшно руско разширение към Индия се оказват напразни. Като компенсация, вицекралете успяват трайно да неутрализират опитите на руснаците да се настанят в Тибет. Така към 1905г., когато Русия и Великобритания се сближават в рамките на Антантата, в Централна Евразия е постигната патова ситуация, допълнена от разпределените зони на влияние в Персия.
Първата Световна война донася значителни изменения в разпределението на силите в Централна Евразия. Рухването на Османската империя след 1920г. Води до обособяването на свободни зони в Сирия. Палестина, Месопотамия и Кавказ. Великобритания успява да си осигури по-голямата част от тези територии, които са й предадени под формата на мандати от новосъздаденото Общество на Народите (ОН). Единствено дневните територии на Сирия и Ливан минават под френски контрол. Ситуацията в Кавказ е малко по-различна. Там грузинци, арменци и азербайджанци се опитват да се възползват не само от разпадането на Османската империя, но и от гражданската война в Русия. Когато след 1919г. Червената армия постига успех над „белите“ царисти, опитите на кавказките републики да запазят независимостта си са осуетени от успехите на комунистите. Впрочем същото се случва и в земите между Каспийско и Аралско море. В крайна сметка, към 1923г. Съветският съюз поставя всички централно-евразийски територии на Руската империя в свои ръце. Великобритания също си осигурява стабилност в своите нови владения, но както се оказва в последствие, локалните процеси, задвижени от идеите на национализма и панарабизма остават неподвластни на имперските сили.
Къде останаха „местните“?
Както видяхме от краткия исторически екскурз, Централна Евразия не престава да е арена на имперско противопоставяне в продължение на повече от 400 години. Османци, перси, руснаци, китайци и англичани си оспорват върховенството на региона посредством директен контрол или налагане на васална зависимост на местните държави и племена. Противно на масовата представа, местните държави и племена не са пасивни участници в този процес. Както вече посочихме в уводната част, местните елити, владетели и кланови вождове се опитват всячески да избегнат поставянето си под зависимостта на големите империи. За целта те използват всякакви дипломатически и военни комбинации, които понякога изглеждат нелогични от дистанцията на времето, но в момента си на приложение отразяват оптималната възможност на различните гео-политически играчи.
Основната част от населението на Централна Евразия се състои от номадски или полу-номадски общества, чиято социална и политическа структура се базира на децентализацията. Клановите структури се подчиняват на по-строги вътрешни правила, но връзката между отделните кланове в едно племе е доста свободна, което често води до флуидност на верността към племенните вождове. Някои от тези общности успяват с времето да кристализират в по-сериозни структури, в които се оформят различни форми на придворен живот и мобилна администрация, която носи някои от чертите на своя еквивалент при уседналите народи, но се отличава на базата на социалните и културни характеристики на номадите. Забележете, не бива веднага да поставяме оценки като „назадничави“ или „изостанали“. Тук терминът „различни“ най-добре обрисува реалното положение на нещата. Това, което тази статия оспорва, е поставянето на етикети за изостаналост и неадекватност. Факт е, че номадските и полу-номадските племенни и държавни структури често съществуват стабилни в продължение на повече от 200 години, постижение с което доста държави в Европа не могат да се похвалят. В такъв случай, редно ли е да твърдим, че тяхната структура е „по-лоша“ или „по-изостанала“ от „европейските“. Същото се отнася и до маниера за водене на война, който тези народи следват. Факт е, че народите на Централна Евразия по-трудно усвояват огнестрелното оръжие. До скоро това се тълкуване от „западната“ историография като принцип на изостаналост. Малко по-задълбочено разглеждане на проблема ни показва, че тромавите, тежки и бавно-стрелящи мускети, използвани от европейците до средата на XIX в, са напълно непригодни за употреба от конски гръб. В това се убеждават самите „западняци“ още през XVII в., когато по време на Тридесетгодишната война експериментите с конница, въоръжена с мускети и карабини се оказват неплодотворни. Едва след техническата революция в средата на XIX в., когато патронът заменя сачмите, а пушките с нарез и самозареждащ се механизъм изместват зареждащите се през дулото мускети, използването на огнестрелно оръжие от конски гръб става практично. Тези оръжия бързо са въведени в кавалерийските части на имперските сили, но тяхната изработка и закупуване са отвъд възможностите на степните народи, които са лишени от постоянни индустриални средища и богата финансова база заради естеството на географските и социални фактори, които доминират в Централна Евразия. В този смисъл, въздействието, което климата и географията оказват върху развитието на един народ не бива да се подценява и би следвало да се включва във всяко сериозно обяснение за техническото и социо-политическото състояние на един народ във всяка една епоха.
Трудно ще е да проследим в детайл всички конфликти, в които местното население взема страната на тази или онази имперска сила с цел да балансира влиянието им в съответната част на Централна Евразия. Въпреки това, можем да установим определени модели, по които действат някои от по-големите племена и ханства. На първо място не може да не започнем с най-стабилната и сериозна „номадска“ държава в Централна Евразия – кримското ханство. То е създадено някъде в средата на XV в., в хода на разпадане на Златната Орда от династията на Гираите, които са по-малък клон на чингизидската династия, която властвала в Златната Орда след отпадането й от големия Монголски улус. Кримското ханство започва съществуването си в един бурен период на конфликти със всичките си съседи. Казанското и Астраханското ханство оспорват претенциите на Гираите за наследство на Златната Орда, докато от запад Литва, подпомагана от Полша започва да се разширява в източна Украйна. Кримското ханство успява да се съхрани в тази враждебна среда и дори преминава в контра настъпление в няколко случая. Въпреки това, опонентите са прекалено много и Гираите правилно оценяват нуждата от повече съюзници и покровители. За тяхно щастие, такъв се явява в лицето на засилващата се Османска империя. След като флотът на Мехмед II Фатих (1446-49;1451-1481) завладява генуезките пристанища в Крим през 1470-те, Гираите решават да дадат васалната си вярност на султана в замяна на което получават военна подкрепа от Истанбул. Османците изпращат поредица от експедиции, които подчиняват Бесарабия и Йедисан, отблъсквайки литовските домогвания. През следващите 300 години кримските татари ще останат най-верният съюзник на османците, а след като Русия завладява ханството им през 1783., доста от тях се изселват и са преселени от османските власт в днешна Добруджа. Но защо става така? Защо, за разлика от толкова други народи на Централна Евразия, татарите от Крим никога не предават своите господари? Обяснението всъщност е много просто – Черно море. Да, правилно, причината е географска. Кримското ханство е уникално с това, че за разлика от останалите племенни и държавни образования, съществуващи в периферията на империите, то си има море, което да го отделя от империята-съюзник. По този начин татарите нямат пряк досег с османците и между двете държави не съществува конфликт на интереси. Нещо повече, османците съвсем целенасочено поддържат автономията на кримските татари и ги насърчават да водят активна външна политика. По този начин османските търговски интереси остават защитени, а големите пристанища Кафа и Кереч са предпазени от чужди нашествия.
За разлика от татарите, останалите държави и племена в Централна Евразия не могат да се похвалят с подобни добри отношения със своите сюзерени. Народите в Кавказ, например, постоянно лавирали между Османската империя и Персия, а след това се обърнали и към Русия, когато и тя се включила в надпреварата. Народите, населяващи земите между Каспийско и Аралско море освен че враждували помежду си, били притиснати от агресивната политика на Персия, Русия и Китай. Това се усетило особено силно през 1740-те години, когато и трите империи били в настъпление за сметка на пограничните народи. Въпреки това казахи, узбеки, ногаи и чагатаи съумели да отбият напора на имперските сили и едва след 1820г. Русия успяла да направи решителен пробив и да се разшири, покорявайки целия регион, заемащ древните провинции Бактрия и Трансоксиана. Както вече споменахме, единствено афганите успели да се изплъзнат от натиска на разширяващите се империи. Това се дължало колкото на упоритостта на афганите, толкова и на географските особености на тяхната родина. Както съвременната практика показва, планинските ридове на Афганистан са сериозно препятствие за логистиката и на най-модерните армии. Можем само да гадаем какви усилия са изисквали експедициите в тези територии през XIX в., а още повече – през XVIII-ти. Всъщност, Афганистан е покорен за последно от Надир Шах през 1738г., и то само защото половината афгани вече се биели на негова страна. Преди това афганите се намирали в полу-васална зависимост от Моголите, които използвали планинските конници по време на своите кампании в Бенгал и платата Декан. След смъртта на Аурангзеб Аламгир (1658-1707г.), властта на моголите постепенно отслабнала. През 1721г. Именно афганите гилзаи донесли гибелта на Сафавидската империя. Две десетилетия по-късно, след смъртта на Надир, Афганистан се превърнал в независимо кралство, където владетелите и племенните вождове подели постоянна борба, така характерна за центъра и перифериите.
Въпреки крайния успех на имперските сили, пограничните държави и племена устояли повече от триста години на постоянния натиск на своите съседи. В този смисъл, виталността и практичността на техните модели на управление не бива да се подценява. Въпреки всичко, края на Първата Световна война поставил начало на една нова епоха – на национализма и суверенните държави, поддържана от устава на ОН и стремежите на всеки народ да се сдобие със своя държава. Този стремеж заменил пограничното упорство и съпротива и въоръжил народите на Централна Евразия с ново средство за борба. Триумфът на империите се оказал мимолетен.
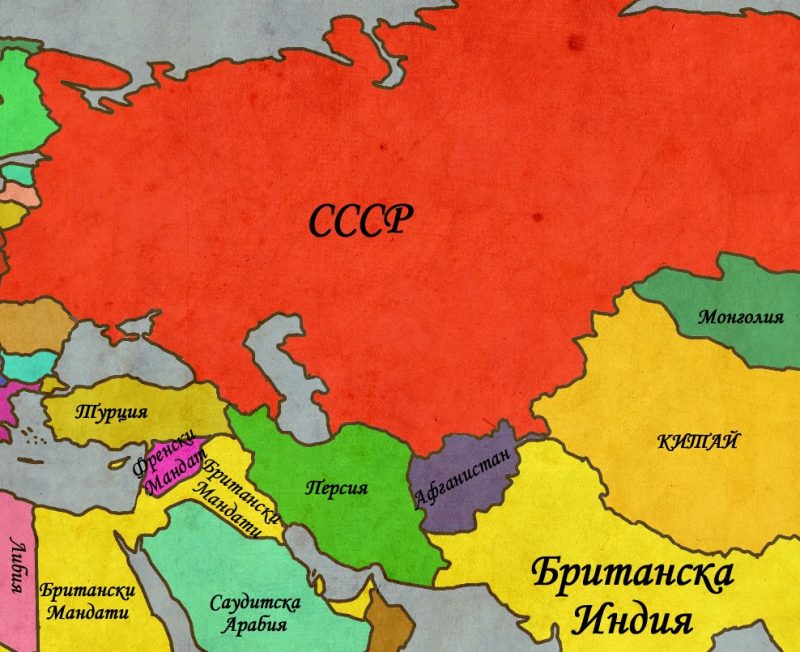
Възход и падение на Великите Сили 1923 – 2012г.
1923г. бележи няколко важни етапа от развитието на международните отношения. На първо място, Съветският съюз окончателно се утвърдил като наследник на Руската империя. Паралелно с това започват преговори за сътрудничество между Съветите и Ваймарската република. Пак по това време в „Близкия Изток“ арабите подемат борбата си за отхвърляне на британското и френско колониално присъствие, а евреи-преселници от Русия строят масово първите кибуци в Палестина. Едно е ясно – решенията на Версайските договори не удовлетворяват нито победителите, нито победените и доста хора търсят реванш.
Последните 90 години са изпълнени с толкова много събития, че спокойно могат да съперничат на изминалите 900. Ето защо отново ще предложим един по-повърхностен прочит на събитията. Целта в случая не е да анализираме изхода от множеството войни или дипломатически конференции, нито да проследим в детайл зараждането на различните нелегални организации, които променят облика на Централна Евразия. Идеята е да се опитаме да отгатнем как става така, че след около век под властта на империите, народите на Централна Евразия намират начин не само да се освободят (повече или по-малко) от опеката на големите, но дори и да контраатакуват по неподозиран начин.
На първо място, всичко се корени в надценяването на Великите сили. Както доста от съвременните проучвания показват, Първата Световна война сериозно изцежда ресурсите на част от страните победителки (най-вече Великобритания и Франция) и отваря мястото за издигане на нови и амбициозни сили (Япония и Италия). САЩ остават сравнително незасегнати, а военно-временните нужди служат като страхотен стимул за презокеанската икономика. СССР от своя страна започва да гради всичките си политически и икономически структури базирайки се на доста заемки от имперския период, но изменяйки ги по начин, който заблуждава за висока доза оригиналност.
Световната икономическа криза от 1929г. позната като Голямата депресия отвлича вниманието на Великите Сили и на обществата като цяло от продължаващите процеси, свързани с национализма и сепаратизма. Народите на централна Евразия започват да търсят път за осъществяване на своите цели. Докато в Индия Ганди призовава за безкръвна революция на мирните протести, арабите започват да формират своите тайни организации, чиято цел е да се преборят не само с присъствието на империите в региона, но и да се противопоставят на еврейските заселници и техните собствени тайни организации, сред които на преден план излиза „Агана“ . Британските власти не остават слепи за тези движения, но империята им е прекалено отслабена за да може да действа активно. Вместо това службите на Негово Величество залагат на тактиката „разделяй и владей“ като подпомагат едновременно и араби и евреи и същевременно извършват публични арести и от двете страни с цел да замажат положението. За щастие на британците, до Втората Световна война нито една друга велика сила няма особени интереси в региона (освен може би Франция) и английската политика успява поне на повърхността да уталожи нещата. Това което британците не осъзнават, е че евреи и араби ги използват също толкова ловко, колкото самите те са насочвани едни срещу други. Субсидиите и оръжието се взимат на драго сърце, но връщаната в замяна преданост е само грим, предназначен да скрие истинското лице на тайните организации и техните цели. Междувременно, СССР продължава да затяга контрола си над своята зона на влияние. Новият съветски вожд – Сталин започва програма на брутално разместване на цели етнически групи в различните краища на централна Евразия. Милиони губят не само домовете но и живота си, а истинските мащаби на този подмолен геноцид тепърва чакат своето разкритие. Паралелно с разместванията, започват строежите на поредица от секретни градове, в които съветската научна мисъл разработва всички сфери на техническия прогрес. Освен това, териториите на т.нар. Среден Изток се превръщат и в удобни места за настаняване на политически затворници, макар Сибир да си остава предпочитаната дестинация.
Втората Световна война приковава вниманието на цялото човечество, но след като изхода й става ясен към 1943г., започват разгорещени дебати за това как да се преразпредели света след това. Ялтенската конференция е повратна точка в този процес, който вече е започнал с окупирането на Иран от съветските и британски сили с цел „превенция от евентуална фашистка атака към нефтените полета на Персийския залив“. След 1944г. Дебатът се фокусира все повече върху Европа и Далечния Изток и Централна Евразия остава някак в периферията на разговорите. Едва след подписването на Парижкия мир през 1947г. нещата се променят. Тук се налага да направим малко отклонение.
Втората Световна война носи една много важна промяна за света. Тя ликвидира концепцията за Концерт на Великите сили и налага двуполюсния модел като водеща гео-политическа матрица за следващите 50 години. Британската и Френската колониални империи, макар да излизат от войната като победители, започват бързо да вървят към своята гибел. Причината? По време на самата война двете страни фокусират всичките си ресурси в Европа и Северна Африка, което означава че останалите им колонии са оставени без сериозен контрол. Нещо повече, част от тях попадат под ударите на Оста (като например Френски Индокитай, Алжир и Тунис), а други са принудени да поемат голяма част от военно-временното бреме (като например Индия, Канада и Австралия). Съответно, тези територии настояват за политически компенсации, голяма част от които на първо време са им отказани. Това води до изостряне на отношенията между империите и част от техните колонии и ускорява процесите на деколонизация. До 1960г. Почти всички колониални владения на Великобритания и Франция се превръщат в самостоятелни държави. В някои случаи, като например Виетнам, се стига до кръвопролитни конфликти. Този процес подминава САЩ И СССР, поради простата причина, че и двете страни нямат колонии и доминиони, а упражняват директен контрол върху всяка територия под тяхно владение. Създалият се политически вакуум трябва да бъде запълнен и по логиката на гео-политическата инерция, двете „свръх сили“ се заемат с тази нелека задача. Първата точка на конфронтация става Европа, където Сталин настоява за своя лъвски пай. В крайна сметка получава каквото иска – 2/3 от Европа са затворени зад „Желязната завеса“. След като Гърция и Турция са предадени на Съюзниците, САЩ и СССР се преместват далеч на изток към Корея, за да уредят поредния спор. В крайна сметка до решение се стига едва през 1959г., с края на Корейската война, която води до изкуственото разделяне на един народ, живял в единство повече от 2000 години. Следващата стъпка е Виетнам, където след две десетилетия на конфликти, Виетконг се налага над „демократичния“ режим, поддържан от САЩ и Франция. По същото време Мао Дзе Дун поставя основите на КНР, с което запечатва Далечния Изток като твърдина на комунизма.
Но да се върнем към Централна Евразия. След изтеглянето на британците, там се случват две основни неща. Първо, гузната съвест на „европейците“ предизвиква създаването на Израел. Второ – схемите на панарабизма не се осъществяват и региона е поделен между множество изкуствено създадени държави, които веднага влизат в пререкания помежду си. Опита на британците да наложат монархичен модел на управление, с марионетни владетели се проваля, подобно на цялостната имперска политика на Острова и скоро поредица от преврати поставят арабския свят под властта на трудно предвидими диктаторски режими. Към 1950г. Великобритания вдига ръце от голямата каша, която е забъркала, обръща се гузно към САЩ и заявява че повече не може да контролира своите бивши земи и че САЩ трябва да спаси положението, за да предотврати съветско проникване в региона. Американците надали са били очаровани от това да поемат ролята на надзирател на още една част от света, но „червената заплаха е надвиснала“ и те нямат друг избор. Къде е проблема? Много просто – САЩ се съгласяват да надзирават една част от света, за която си нямат никакво понятие. Американците не разбират местната култура и политически традиции и вярват че сляпото наливане на пари и оръжие е достатъчна грижа за защитата на интересите им в региона. Колкото и странно да изглежда, СССР, който като пряк приемник на Руската империя наследява и вековните й връзки с Централна Евразия, допуска същата грешка. В крайна сметка облагодетелствани са държавите, които ловко маневрират между двата лагера и на практика са на издръжката и на двете супер сили. Въпреки това, САЩ и СССР съвсем не са безсилни в опитите си да наложат контрол. Докато съветите залагат на директните отношения с местните диктаторски режими (не на последно място заради сходствата им със съветския модел), САЩ се ориентират към финансирането на различни „терористични“ организации там, където съветите са спечелили диктаторите или пък директно финансират диктаторските режими, както например става с Египет от времето на Насър.
Тъй като без въоръжени конфликти не може да се мине, САЩ и СССР намират идеалното решение – Израел и арабите. Разбира се, там нещата са доста по-сложни, но без да навлизаме в детайли, можем да обобщим арабо-израелските конфликти като отваряне на голям пазар за съветско и американско оръжие, поддържан както от комунистите в Москва, така и от евреите-капиталисти, които от век и повече контролират американската икономика. Войните разбира се са краткотрайни и ограничени по мащаб, за да не би да прераснат в нещо наистина сериозно. В крайна сметка, народите между Нил и Ефрат се оказват само пешки в големите сделки с оръжие на свръх силите и на техните собствени „правителства“, които не се свенят да платят собствената си състоятелност с кръвта на своите сънародници.
Малко по-различно стоят нещата в Иран. Там САЩ подкрепят диктаторския режим на шаховете, въпреки явните погазвания на доста от човешките права, за които САЩ се бие в гърдите че защитава, и върху които се гради демокрацията отвъд Океана. Но както е казал Франк Хърбърт – „важното е подправката да тече“. В случая „подправката“ е черното злато, което изтича от Персийския залив и захранва световната индустрия, било то военна или цивилна. Надпреварата между САЩ и СССР върви с пълна сила и нито една от двете не може да си позволи несигурност по отношение на доставките на петрол. Както се оказва, скоро и двете остават излъгани в увереността си. През 1973г. Държавите от персийския залив, с цел да притиснат САЩ и съюзниците му заради подкрепата им за Израел в Шестдневната война, увеличават цените на суровия петрол, което на свой ред предизвиква поредната сериозна финансова криза, чийто отражения се усещат като вторични трусове чак до средата на 90-те години на миналия век. Директния удар пада върху икономическото развитие и на двата лагера. Външните дългове, генерирани чрез заеми скачат в пъти, което се отразява толкова тежко на СССР и компания, че след 1985г., краха на комунистическия блок става неизбежен. Франция и Великобритания също са пометени, а икономическите негативи на кризата са причина за падането на доста от правителствата в тези държави чак до 1995г. САЩ излизат сравнително невредими, но получават важен урок за това, че военното превъзходство не означава нищо, когато арабите врътнат кранчето на петрола. Следващата изненада се случва само шест години по-късно, когато в Иран се случва нещо неочаквано – ислямска революция. Идването на шиитското духовенство на власт и премахването на светската монархия се оказва неизгодно колкото за САЩ, толкова и за СССР. Първите губят верен съюзник в региона и си спечелват вечен идеологически враг. Вторите се сблъскат с една система базирана на най-непознатата материя за един комунист – религията. Революцията в Иран води до две незабавни събития – Ирано-Иракската война (1980-1989г.) и съветската инвазия в Афганистан.
От една страна Садам се опитва да отвлече вниманието на изстрадалите си поданици като обяви свещена война срещу шиитските еретици. От друга СССР се опитва да се справи с новосъздадената несигурност като се разшири териториално. САЩ не могат да останат безучастни, особено след като дипломатическата им мисия е била пленена от иранците. Вашингтон подкрепя Ирак чрез оръжие и пари, също както и талибаните в Афганистан. Пак по същото време американците подпомагат Бин Ладен и неговата „Алкайда“ като обучават терористите и ги снабдяват с оръжие и средства за борбата им със съветите. В крайна сметка и двете войни завършват безрезултатно от гледна точка на териториални промени. От гледна точка на бъдещето, резултатите са ключови. Афганистан се превръща в надгробното слово на СССР, поглъщайки последните материални ресурси на червената империя. 1989г. Ще отбележи не само окончателното изтегляне на червената армия от Афганистан, но и началото на края на социалистическия блок. България, Румъния, Полша, Чехословакия и Унгария отпадат от СИВ и Варшавския Договор. Но нещата не спират до тук. Кавказките републики, както и съветските републики между Аралско и Каспийско море отпадат от СССР. Империята, градена от Романви в продължение на 300 години рухва за 3. Триумфът на САЩ изглежда пълен – комунизма е съкрушен, а американските съюзници печелят в региона. Франсис Фокуяма ще заключи, че е дошъл края на историята.
Само че радостта се оказва пресилена. САЩ по детински захвърлят старите си играчки – терористите, забравяйки да си приберат огромните оръжейни арсенали, които са им оставили. Забравили са също да си отбележат, че след като съветската заплаха вече я няма, САЩ ще бъде единствения виновен за политическите несполуки на региона. Съвсем скоро от добър съюзник и кредитор, САЩ се превръща в символ на империализма и империя на злото – термини, които само половин десетилетие по-рано Рейгън е използвал по повод СССР. Първата война в Залива (1991г.) само подкрепя нарастващото недоверие на народите от Централна Евразия към САЩ. Въпреки това успехите на Буш Старши замазват очите не само на обикновения американец, но и на експертите им в Пентагона и Капитолия. Създаването на бази в Централно-азиатските републики и и Грузия се брои за поредния силен удар срещу рухващата съветска империя. И действително, до идването на власт на Путин през 1999г., нещата изглеждат обещаващо за САЩ. Но началото на новото хилядолетие доказва, че радостта е била прибързана. Руската федерация, издигнала се като феникс от пепелта на СССР бързо си връща позициите, благодарение на зависимостта на Европа от руския газ и нефт, който както щедро тече, така и щедро може да бъде спрян по тръбопроводите. Паралелно с това, Русия инициира създаването на ОНД, което на практика привързва всичките й бивши републики без Грузия и Балтийските страни в едни пакт, който представлява по-децентрализиран вариант, лежащ по средата между ЕС и НАТО. Путин приключи кризата в Чечня, а наскоро доказа на Грузия, че когато нещо се случва в задния двор на Русия, другите големи държави си мълчат. Едновременно с тези процеси, отпадането на „червения враг“ даде шанс на Франция, Германия и Великобритания да напуснат директната американска орбита и да започнат да водят по-независима политика, както си пролича по време на втората Война в Залива.
11.09.2001г. се превърна в една от най-противоречивите дати в човешката история. За крайните ислямистки организации тя отбеляза началото на нова ера, за САЩ – началото на Войната с Терора, а за останалия свят се превърна в доказателство, че САЩ не е неуязвима дори на своя територия. Трагедията на всички семейства, засегнати от атентата в Световния търговски център е неминуема и не подлежи на тълкуване. Това което трябва да се тълкува са последвалите събития. САШ побърза да отвърне на удара, за да не изглежда слаба в очите на света. Афганистан беше нападнат и завладян светкавично, последван от Ирак две години по късно. Но това не изненада никой. Едва ли има човек с трезва преценка, който да не признава факта, че САЩ е най-голямата военна сила, която историята познава. В този смисъл, ясно е, че единствено страните в Съвета за Сигурност на ООН могат да бъдат реален опонент на Вашингтон и дори тогава шансовете за успех на САЩ при сблъсък едно в едно са над 70%. Истинските проблеми на американците не са свързани с побеждаването на враговете им в Централна Евразия, а с това как да управляват завзетите земи след победата. И тук на преден план излезе очевидната слабост и липса на подготовка сред експертите във Вашингтон. Липсата на такт и всякакво разбиране на местните схеми на разпределение на властта и осъществяване на контрола център-периферия доведоха до десетилетие на несигурност и непрестанни кървави атентати. Да, САЩ ликвидираха два диктаторски режима…само за да ги заменят с анархия, дестабилизация и липса на адекватна властова политика. Икономиките, социалната сигурност и законовия ред рухнаха в прахта на лаицизма на новите окупатори. Защото колкото и да се бият в гърдите че са носители на свобода и демокрация, американските военни никога няма да бъдат нищо повече от окупатори за местното население, което се е нагледало на различни форми на диктатура за последните 70 години.
И точно когато всичко като че ли беше достатъчно заплетено, Централна Евразия ни изненада с нова гео-политическа бомба, която затвори устата на всички световни експерти по регионалните проблеми – Арабската Пролет. Арабите добре подбраха момента да разрушат реда, наложен им от САЩ и СССР след 1945г. Световната икономическа криза парализира способността на НАТО да води политика, която следваше в началото на новото хилядолетие, а Русия от своя страна е все така заета с вътрешните си проблеми и удържането на приятелските режим в страните, завършващи на „-стан“. Останали без подкрепата на своите големи спонсори, диктаторите започнаха да капят като есенни листа пред зимата на арабското недоволство и последвалата я пролет изпълнена с упорит стремеж към промяна. Новосъздадената ситуация е напълно непредвидима. Тунис и Либия уж извървя пътя към промените, но съдбата им е все така неясна. Египетския народ води отчаяна борба срещу военните за да предотврати издигането на нов Мубарак. Асадова Сирия все още гори в пламъците на гражданското неподчинение, подхранвани от правителствените снаряди. Йемен като че ли притихна, но едва ли. Ливан и Ирак все още съществуват на ръба на държавността, готова всеки момент да рухне. Историята на региона се пише всеки ден и все още е рано за крайни резултати. В така създалата се ситуация, на преден план излизат интересите на новите икономически колоси – Китай, Индия и Бразилия. Трите страни използваха кризата за да се съюзят с Русия и да разбият монопола на западните икономики. Отделно от това, Пекин възобнови интереса си на запад, където оръжието от Китай е не по-малко търсено от своите руски или американски еквиваленти. Непреклонната политика на червените вождове в Поднебесния комунизъм стана явен по време на кризата в Сирия, и изглежда, че Китай тепърва ще разгъва пипалата на своята политика в региона..
Къде останаха местните? Тихомълком от взора на големите сили, Пакистан създаде своя ядрена програма, последван от Изараел, а сега и от Иран. Като добавим Индия и Русия, Централна Евразия се оказва съсредоточната точка на световните ядрени сили. Получената взривоопасна смес заплашва не само локалния но и световния мир и е време големите лидери по-внимателно да премислят ходовете си в региона. Арабите вече са доказвали, че могат да врътнат кранчето на нефта и да слушат своята собствена съвест и интереси. Израел е винаги непредсказуем, поставяйки личните си интереси над всичко. Не бива да се подценява и възможността на народите между Каспийско и Аралско море да отхвърлят руските марионетки, които ги управляват и да поемат по свой собствен път. Кавказ е всичко друго но не и спокоен. Армения и Азербайджан продължават спора си за Нагорни Карабах, докато Осетия, Абхазия и Грузия са вплетени в сложна игра с Русия, която всячески се опитва да избегне втора Чечня. Отново се наблюдава характерната от векове игра на смяна на съюзи и каузи с цел да се тушира влиянието на големите имперски съседи. Мислим си, че технологичните революция и абсолютизирането на държавните системи е променило правилата на играта, но както Централна Евразия доказва, това съвсем не е така. Регионът е един гео-политически кръстопът, чиято значимост не е намаляла и със стотна за последните пет столетия. Заключение
Централна Евразия е една от люлките на човешката цивилизация. Тук било измислено колелото, тук се появили първите градове, първите архиви и първата азбука. Първите империи мерели сили върху древните бойни полета, рухвали и нови се издигали, оставяйки колосални културни и политически традиции, които и до днес доминират начина, по който нещата се случват в региона. Въпреки технологичния напредък на човечеството, географията, климата и традициите се оказват неподатливи на сериозни промени и процесите продължават да се случват в общи линии така, както са се случвали преди петстотин години. Както рухването на Тимуридската империя в края XV в. поставя началото на преразпределение на силите в региона, така и рухването на СССР през 1991г. постави началото на ново разместване на гео-политическите пластове. Стари титани се изправят, предизвикани от млади богове олимпийци, но това, което като че ли забравят във висотата си, е че дребните души са тези, чиято вяра и отдаденост крепи великаните. Забравят също, че тези уж дребни гео-политически пешки лесно могат да сменят боята си в случай че партията шах не върви в тяхна полза. Правили са го преди, правят го сега и ще го правят и за в бъдеще. Природата е отнела на Централна Евразия богатите почви, огромните метални залежи и възможността за създаване на лесни икономически и транспортни коридори. В замяна й е дала непреодолим за външните врагове терен и черното злато – нефта, както и неговата сестра – газта. Местните народи добре са се научили да компенсират недостатъците, извличайки максимална полза от своите активи. Там където западноевразийците прилагат изкуството на войната, централноевразийците са се научили да използват изкуството на дипломацията. Войната не им е чужда и те не са лоши войни, но като че ли търпеливото изчакване, подмолните рокади и смяната на съюзниците са по-подходящи за постигането на дълготрайни резултати. Подобна политика остава недостижима за империите, увлечени в завоевателен устрем. Агресивната и прибързана политика често се разбива в подводните скали на търпеливото изчакване. В крайна сметка, още преди векове Конфуций е заключил, че ако достатъчно дълго и търпеливо седиш на брега на реката, течението ще изхвърли трупа на врага ти. Удряй врага когато е слаб, съюзявай се с него когато е силен, а с враговете му когато е прекалено силен – така може да се обобщи философията на народите на Централна Евразия. Старите империи – Персия, Османската, Моголската и Руската ги няма. След тях изчезнаха Британската, Френската , а след това и Съветската. Сега САЩ се тресе на ръба на финансова криза, ресурсите му – разпилени на десетки фронтове из цял свят, ала най-сериозните са разположени именно в централна Евразия. От друга страна, узбеките все така населяват поречието на Амру Дъя, казахите обитават степите на южен Сибир, а кавказките народи вървят по пътя на своята политическа фрагментарност въпреки опитите на големите да поддържат единение там, където то никога не е съществувало. Афганистан, непокорен през вековете така и не позна пълния американски контрол вече повече от десетилетие, а Ирак, тази изкуствена спойка на британската политика се лута в търсене на една маргинална идентичност, но остава неподчиним за задружните усилия на НАТО и новата местна власт. Центално-евразийците имат навика да стоят и да гледат как империите рухват около тях, точно както и ние българите обичаме да правим същото. Кръстопътното положение предполага досег с множество велики сили, но също така изгражда един особен имунитет към тяхното влияние. Всякакво подчинение остава само на повърхността, докато истинските процеси и стремежи остават дълбоко скрити и непредвидими. След векове на борби, междуособици и конфронтации, Централна Евразия е изпълнена с „жизнени“ местни народи и племена и с надгробните плочи на десетки империи…
ПРИЛОЖЕНИЯ
Централна Евразия в 1400г. Границите на държавите са приблизителни.
Централна Евразия в 1500г. Границите на държавите са приблизителни.
Централна Евразия в 1600г. Границите на държавите са приблизителни.
Централна Евразия в 1700г. Границите на държавите са приблизителни.
Централна Евразия в 1800г. Границите на държавите са приблизителни.
Централна Евразия в 1900г. Границите на държавите са приблизителни.
Централна Евразия в 1923г.
Централна Евразия в 2012г.
Конфликтни зони в Централна Евразия към януари, 2012 г.
ПРЕПОРЪЧИТЕЛНА ЛИТЕРАТУРА
На Български:
Даймънд, Джаред – Пушки, вируси и стомана, София, 2006г.
Даймънд, Джаред – Колапсът, София, 2007г.
Кенеди, Пол – Възход и Падение на Великите Сили, София 2011г.
Кетъруд, Кристофър – Кратка история на Близкия Изток: От Авраам до Арафат, София, 2009г.
Кисиндджър, Хенри – Дипломацията, София, 2009г.
Мантран, Робер – История на Османската Империя,
За съдържанието на термина „властови център“ виж по-долу.
Наричани най-често от нас българите – „Западна Европа“
Въпросът за това дали САЩ е империя или не е много много спорен, но е факт, че ако не институционално, то поне доктринално поведението на САЩ определено е имперско. Това, разбира се, е закономерно предвид размера и възможностите на тази държава. Бих бил повече от щастлив ако тази статия се окаже стъпка напред в развенчаването на мита, че само държави, управлявани от император са империи.
Да не се бърка с Монголската Империя. Моголите (Мугали, Мугули) са потомци на армиите, подчинени на Чингиз Хан, които се заселили в централна Евразия и по-специално в Афганистан. Империята на Моголите е създадена от Захир ад-Дин Мухамад Бабур, владетел на Кабул и потомък на Тамерлан през 1526г. След като узбеките завладяват Самарканд, Бабур решава да насочи силите си към Северна Индия, където Делхийският султанат бавно се разпада. След битката при Панирпат (1526г.), Бабур подчинява целият Делхийски Султанат. Потомците му Акбар и Аурангзеб ще разширят империята на юг и изток и през 1700г. Империята на Моголите обхващала по-голямата част от днешна Индия. Империята е ликвидирана през 1858г., когато земите й окончателно стават част от Британската империя.
Виж приложените карти които показват изменението на пограничните зони между 1400 и 1900г.
Според историографската традиция, Ертогрул (1230-1281г.) и Осман (1281-1324г.) са първите бейове (бегове) на Османците, които завладяват малка територия в Северозападна Мала Азия, която се превръща в ядрото на Османската империя.
Термин, популярен в англоезичната литература, обозначаващ комплекс от социални и културни параметри, които определят характеристиките на дадено общество.
Виж картата за 1500г. в Приложението.
Федерацията на Белите Овни се появява като локална сила в началото на XV в. Като следствие от завоеванията на Тамеран (Тимур Ленг). Основният опонент на Аккоюнлу е другата туркменска федерация – Каракоюнлу (Федерация на Черните Овни) В продължение на половин век двете страни водят постоянни конфликти в северна Месопотамия и западен Иран. В крайна сметка султанът на Аккоюнлу – Узун Хасан (1453-1478г.) надделява над своите опоненти, завладява Каракоюнлу и разширява държавата си, която в зенита си обхваща днешните Ирак, Кувейт, Армения, Азербайджан, Иран, части от Турция и Сирия.
Империята на Тимуридите е създадена от Тамерлан (Тимур Ленг) (1370-1405г.) в следствие на поредица от кампании, които покриват земите от Делхи до Измир и от Волга до Персийския залив. Тимуридската империя изживява кратък разцвет през първата половина на XV в., но след смъртта на Шакрух (1405-1448) и Углуг Бек (1449-1450), империята се разпада на множество ханства, всяко от което претендира за върховенство.
Мухамад Шайбани (1500-1510) – владетел на Узбеките, който за кратко управлява Трансоксиана и Североизточен Иран. Загива в борба със шах Исмаил. Владенията му са наследени от чичовците му. Узбеките остават значителна сила в земите между Каспийско и Аралско море до средата на XVIIв., когато мястото им е заето от Казахите и Чагатаите.
Бабур е прякор, който води произхода си от персийската дума бабр (тигър)
Делхийският султанат е мюсюлманска държава, разположена в централните части на Северна Индия със столица Делхи. Издига се през Късното Среновековие като водеща военно-политическа сила на Индийския субконтинент. През 1399г. Тамерлан превзема Делхи и опустошава града, но въпреки този тежък удар султаната се съвзема под управлението на династията Лоди. Към края на XV в. Политиката на династията настройва местните елити срещу тях. Победата на Бабур при Панирпат е последният удар, който слага край на вече разклатената власт на династията Лоди.
Династията на Исмаил, която ще управлява Персия до 1721г.
Сафавидите са шиити а Моголите – сунити.
Двете империи си оспорват властта над Афганистан.
Голяма източноевропейска държава, която представлява персонална уния между кралство Полша и Великото херцогство Литва. Двете държави са династични свързани от началото на XV в., а унията става официална с договора от Люблин (1569г.). Посполитажеч (или Жечпосполита) обхваща териториите на днешните Полша, Литва, Латвия, Беларус и Украйна. Макар и да е една от големите сили на своето време, мощта на Посполитажеч запада заради вътрешните конфликти между изборните крале и големите магнати (шляхта). След 1700г. Посполитажеч започва да губи големи дялове от територията си за сметка на Русия, а по-късно и на Австрия и Прусия. През 1795г. се случва последното от Трите Деления на Посполитажеч, чиято територия е изцяло поделена между Русия, Прусия и Австрия.
Монголската династия Юан (1271-1368) управлява Китай от разпадането на монголската империя до бунта на Минг. По времето на династията Юан Китай достига най-големия си териториален размер, съчетан с отворена икономика и силно чуждестранно влияние. Последните два фактора сериозно допринасят за последвалата затвореност по времето на империята Минг.
Императорът Йонгле (1402-1424г.) е брат на първият император от династията Минг – Хонгву. Светското му име е Чу Ди и до 1402г. Служи като управител на Пекин. След като организира заговор, Чу Ди сваля своя племенник и сам заема трона, като мести столицата от Нандзин обратно в Пекин и построява прочутия „Забранен Град“.
Система от валове, ровове и фортове, които пресичат откритите равнини на Русия. За първи път подобна система от укрепления е използвана срещу татарите през XIV-XV в., в областта Тула. По-късно с изместване на границата на юг, най-значителната „засечна черта“ става Белгородската. През XVIII в. подобни укрепления се появяват и източно от река Урал за да защитават новите руски владения от Казахите, узбеките и другите племена, населяващи земите между Каспийско и Аралско море.
Калмиките са най-силното не-монголско племе в Централна Евразия. Силата им нараства след 1600г., когато постепенно се обединяват в нещо като конфедерация от кланове със свой хан. Калмиките започват да воюват със всички племена от монголското семейство, което им печели прозвището „страшилище за монголите“. Москва предвидливо си осигурява тяхната васална зависимост и често ги използва във войните си с ногаите и кримските татари. Калмиките играят важна роля в кампаниите не Петър I в северното Причерноморие и Кавказ.
Тази практика достига своя разцвет по време на Първата Световна Война.
Британската зона на влияние включва областта Балуджистан и пролива Ормуз, докато руската зона се простира по южното крайбрежие на Каспийско море.
Този казус е подробно разгледан в книгите „Пушки, вируси и стомана“ и „Колапсът“
Единствената граница между двете държави е по кримското крайбрежие на Черно море, където османците контролират трите основни пристанища – Кафа, Кереч и Азак (Азов).
Наричан „Последния велик завоевател на Азия“, Надир (1736-1747г.) се издигнал по времето на разпадането на Сафавидската империя (1721-1728г.) Надир събрал огромна армия с която обединил Иран, завзел Афганистан, Оман и Кавказ и изтласкал османците от Месопотамия. След смъртта му империята се разпаднала, тъй като разходите, генерирани от кампаниите на Надир се оказали непосилни за хазната.
Моголски император, син на Шах Джахан и Мунтаз Махал. По негово време Моголската империя достига своя зенит.
Агана е една от първите тайни организации на евреите в Палестина. От редовете й произлизат повечето генерали на израелската армия след 1948г. Агана се превръща в базата, върху която се изграждат израелските сили за сигурност.
През 609 г. пророк Мохамед, неговата жена Хадиджа, неговият слуга, един от братовчедите му и техният приятел Абу Бакр създават една нова религия в безлюдното и затънтено градче Мека в Арабия. Религиозната организация, която през 740 г. ще се разпростре от бреговете на Испания до Индия и границите на Китай, през 613 г. присъединява в своите редици още 14 съмишленици. През тази година Мохамед се провъзгласява за пророк пред съгражданите си.
Хегира
През следващите десет години десетки от тях се присъединяват към каузата му, но останалите са повече и подгонен от техния гняв, през 622 г. той е принуден да напусне Мека. Това е първата година на „бягството“ (хегира) – първата от летоброенето на исляма. Мохамед се завръща в Мека като победител през 630 г. Умира там през 632 г.
Сунити
Също както двете най-големи християнски църкви могат да проследят своята история до оригиналната, основана от Христос, така и двете главни секти в исляма претендират да са основани от самият Пророк Мохамед. Схизмата, разделила шиити и сунити и дала им идентичност на различни секти, започва през 632 г. -само няколко седмици след смъртта на Пророка и още преди да са обособени някакви теологични различия между тях. Сунитите, или „традиционалистите“, базират своята вяра на Корана и традиционното описание на учението и живота на Мохамед. Те съставят основното течение в ислямската вяра. Те разпознават и поддържат халиф Абу Бакр – първия наследник на политическия пост на Мохамед, като глава на Ислямската държава.
Шиити
Шиитите поддържат, че братовчедът и племенник на Мохамед Али ибн Аби Талих е законният наследник на Мохамед. Причините са в това, че той е най-близкият роднина по мъжка линия на пророка. Освен това Мохамед дал знак за това, като държал Али далеч от делата си като пророк. През 655 г. Али е издигнат до поста халиф, но той е едва четвъртият поред мъж, заел този пост. Според шиитите той е първият, а всички преди него са били узурпатори. Шиитизмът е дело преди всичко на поддръжниците на Али, а не на него самия, в основите му е убеждението в заслугите на фамилията и в божествения произход на правото на една фамилия да управлява по наследство. Развили се по-късно и независимо, теоретичните основи на сунитите и шиитите откриват различия и по ред други въпроси.
Исмаили
Исмаилизмът е неортодоксално разклонение от шиитите, председателствано понастоящем от Ага Хан. То започва като група нелегални и независими движения в Сирия, Ирак и Арабия през късния VIII век. Те се сближават някъде около 800 г., приемайки Ислямия, син на Джафар ал Садик за седми имам (духовен водач) на шиитите. Теза, която се споделя от останалите шиити. Тяхната най-забележителна черта е осемстепенната система за обучение, пропагандирана от техните мисионери. Първата степен е общоприета религия – за обикновените вярващи. По това те не се различават от сунитите. По-високите степени включват отказ от религията и алегоричен прочит на Корана. В тези степени исмаилите възприемат свободното мислене, светската философия и материалистичен анализ на Вселената.
Учените мистици в ислямския свят са известни като суфи от съзерцателната значимост на звуците S, U и Р. Те почитат значими личности от древността, като Питагор и Христос, и ги наричат „суфи“-учители. В действителност тази секта е свързана с трудовете на големите мистици от IX век, живели в Ирак и Египет. В частност ал Мухашиби, Абу-Джазид ал-Бистами, ал-Халя и Дбу- Нин ал-Масри поставят основите на суфи мъдростта – отричаща задълбоченото учене и заела се да води своите последователи до действителната истина.
Дервиши
Суфите предлагат различни практически пътища към знанието, което възхваляват. Повечето включват в себе си различни форми за самодисциплина и самоотричане от предимствата на словото. В началото на IX век последователите на един от тези съвети сформират различни ордени на дервишите.
Бистами дервиши
Най-стар от тези ордени е на Бистами дервишите, сформиран през 874 г., за да следва учението на Абу-Джазид ал-Бистами, починал през следващата година.
Въртящи се дервиши
Най-известен от всички ордени е Мевлеви, основан от Джалил ал-Дин Рюми (1207 – 1273), известен поет от Централна Азия. Рюми е забелязан и обучен от дервиша Шам ал-Дин от Табрус, докато двамата живеят в Коня – Турция. Скоро след смъртта на последния, през 1244, Рюми основава ордена, известен като „Въртящи се дервиши“. При церемонията sema те се завъртат бързо около лявата си пета, като държат лактите си широко разтворени. Турското правителство забранява сектата през 1925 г. В музея „Мевлеви“ в Коня може да бъде видян електрически модел на въртящ се дервиш.
Съвременните християни завършват своите молитви с „амин“. Те използват тази може би най-древна дума в нашия речник, за да помолят за одобрението на единствения истински Бог. „Амин“ е термин, използван от египтяните, първите монотеисти, още през IV хилядолетие преди Хр., за да обозначат с него единствения Бог. Думата и религията са заимствани от евреите по време на тяхното пребиваване в Египет. Египтяните не са искрени, когато казват, че не знаят истинското име на Бог. По време на богослуженията те го крият зад много други и различни имена.
„Той превзема всички сърца и ги отнася надалече.
Любовта към тебе прави ръцете да се отпуснат,
твоите хубави дела правят ръцете ти да треперят.
Всички сърца се разтапят, когато те видят. O, човешка фигура!
Един, създател на всички неща. O, един-единствен,
Творец на всичко, кой си?“
Това са думи от един монотеистичен химн, създаден от около второто хилядолетие пр. Хр. Други не толкова добре запазени документи ясно показват, че вярата в единствения и върховен бог е била постигната още през четвъртото хилядолетие пр. Хр. Множеството младши богове и богини са тачени от тези вярващи само като създадени от върховното божество творения с делегирана сила.
Атеизъм
Първият философ, в чиито теории е включено категоричното отхвърляне на божествата, е грък – Диагорас от Мелос. Той е ученик на великия Демокрит от Абдера. Теорията за атомния строеж и научното разбиране за съдбата, създадени от Демокрит, окуражават Диагорас да разработи атеизма. Той преподава своята теория в Атина. Обвинен в безбожие, Диагорас е принуден да напусне града, но продължава делото си в Коринт.
Агностицизъм
„Относно боговете аз не мога да кажа дали те съществуват или не, или на какво приличат. Към това има много пречки за познанието. Неяснота на самите въпроси и ограничеността на човешкия живот.“
Това е съждение на гръцкия философ Протагор от Абдера (485 – 415 г. пр. Хр.), повече известен с догмата „Човек е мярката за всички неща“. Протагор е първият „софист“ – един от групата учители, които искат да се плаща такса и по тази причина си навличат презрението на Сократ и последователите му. Това е единствената причина Протагор да не е толкова известен.
Сикхи
През XV век Индия е разкъсвана между настъпващото, проповядващо равенство и единствен бог мюсюлманство и хинду религията, защитаваща кастовото разделение на обществото. Това окуражава някои учители да се опитат да разчупят религиозните бариери. Един от тях е Нанак (1469 – 1539) – по рождение от общността на Хунду. Той обикаля из Индия и проповядва, че всички церемонии и книги на различните религии не са от значение за същината/силата на отделния човек. Той отрича обосноваността на кастовата система и провъзгласява единствено върховния Бог, достоен за обожаване от вярващите от всички религии. Неговите поеми са пълни с пренебрежение към частните особености на исляма, хинду и будизма, но преклонението пред Бог е винаги в тях.
Ако всички хора живели някога в миналото, бяха живи сега, за да опишат целия Му път, той пак няма да бъде описан пълно.
Поемите на Нанак сочат на всички вярващи един хуманен и приложим път за преодоляване на блатото на религиозните различия. Но историята си прави шега с неговото дело. Вместо да бъдат заличени религиозните различия, се ражда нова религия, която влиза в остър конфликт с хинду.
Гуру
Сикхите се възприемат като отделна и ясно обособена общност с един водач – „гуру“ (учител), и разпознават Нанак като свой първи гуру.
Общ произход на живота
Друга изключително важна идея, която може да се намери в сборника „Хуан Нан Дзъ“ (II в. пр. Хр.), е, че всички съществуващи на Земята живи същества имат един общ праотец. Това е виждането на един от авторите, който сравнява сътворението с дърво – ствол, от който излизат безброй клони. В тази концепция се подразбира, че разликите между животинските видове прогресивно се увеличават с поколенията, които ни отделят от общия прародител на всички форми на живот.
Манихейство
Подобно на зороастрийците манихейците вярват, че светът е в ръцете на конкуриращите се сили на светлината и мрака и борбата между тях няма начало и край. Те настояват да сме милостиви и да пазим растенията, огъня, водата и особено животните. Тази религия е основана от Мани (роден в Персия през 240 г.). Мани описва себе си като следващия вестител на Бог – след Буда, Заратустра и Исус. Неговата вяра съчетава персийски, средиземноморски и индийски религиозни концепции.
Джайна
Тази религия е създадена през VI в. пр. Хр. от съвременника и сънародника на Буда Вардхамана Махавира Тиртанхара от Магада в Североизточна Индия. Тя проповядва, че всеки човек, животно, растение и дори камък са надарени със своя индивидуална душа. Джайна отрича съществуването на върховно божество или творец. Най-забележителната догма на Махавира е, че Бог е само последна и висша проява на силата, присъща на човешкия дух.
Гръцкият философ Емпедокъл (495 – 435 г. пр. Хр.) от Акрага в Сицилия в съчинението си „Върху природата“ описва това, което сам нарича „космически кръговрат“. Цялото съдържание на Вселената някога е било събрано в хомогенна сфера. Впоследствие то се разлетяло на всички страни, отнесено от вихрушка, и постепенно достигнало до формите на Вселената, в която живеем. В края на краищата всичко ще тръгне в обратната посока и отново ще се свие в огромна сфера, за да се разпръсне отново в един процес, който се повтаря до безкрайност.
Безкрайност на Вселената
Съгласно Анаксимандър (610 – 540 г. пр. Хр.), гръцки философ, живял в Милет (в днешна Турция), Вселената е безкрайна и включва нашия собствен свят и безброй други подобни на него.
Безкрайност на времето
Същият философ описва безкрайността като процес на непрекъснато съзидание и рушене на Вселената, вярвайки в безконечността на времето.
Безкрайност на материята
Отначало нашата Вселена е била възприемана като заемаща ограничено пространство със Земята в центъра (или на дъното) и звезди, обикалящи по затворени орбити по границите му. Анаксагор от Клазомена (500 – 429 г. пр. Хр.), гръцки философ, работил в Атина, развива теорията, според която нещата са безбройни, което означава, че Вселената няма граници.
E = m.c2
Обединението на тези два фундаментални принципа се съдържа в равенството E = m.c2 (енергията е равна на масата по квадрата на скоростта на светлината). Откритието е направено през 1907 г. от Алберт Айнщайн, бащата на теорията на относителността
Запазване на материята
Принципът, според който материята не може да бъде създадена или унищожена, е въведен за пръв път от гръцкия философ Емпедокъл от Акрага в Сицилия. В своето съчинение „Върху природата“ (V в. пр. Хр.), където въвежда естествения подбор и „Теорията за големия взрив“, той постулира, че всичко във Вселената е вечно, че раждането и смъртта са само думи, които ние използваме, за да опишем преобразуването на нерушимата материя. Този принцип е приет веднага от създателите на теорията за атомния строеж, включително и от Демокрит. Те разбират, че всичко, възприемано като осезаема материя (като камъните и езерата), в действителност е струпване на невъобразимо малки частички, които, движейки се, създават илюзията за обем и вещественост. Така те развиват една по-прецизна версия на принципа за запазване на материята, а именно, че тези частички са вечните (а не непрекъснатата материя, която ние наблюдаваме) и че те не се раждат, нито умират. В по-късни времена последователите на Теорията за големия взрив изтъкват принципа за запазване на материята срещу Теорията за равновесието, в която се предполага непрекъснато образуване на нова материя.
Запазване на енергията
Подобно твърдение, но за енергията – че тя не може да бъде създадена или унищожена – е възприето едва след като сър Исак Нютон (1642-1727) го дава на света в своята „Philosopiae Naturalis Principia Mathematica“ през 1687 г. В ръцете на неговите последователи този принцип е превърнат в крайъгълен камък на физиката.
Отричане на причинността
Дискретността на времето е използвана по-късно с цел отричане на днешната представа за действителна причинно-следствена връзка, базирана на представата за непрекъснат ход на времето. Според Абу Ишак ал-Назам – арабски теоретик на Исляма, живял през IX в. – Вселената във всеки отделен момент се преподрежда от Бог чрез непрекъсната промяна на положението на атомите. Според него и неговите последователи от сунистката школ природните закони, открити от науката, са просто навиците на Бог при преподреждането. Чудесата възникват тогава, когато Бог се отклони от навиците си и създава явления, които не са в съответствие с тези „научни закони“. Дискредитирането на времето и отказът от причинно-следствена връзка са тясно свързани с квантовата физика.
Дискретност на времето
Ние възприемаме времето като нещо непрекъснато, но точно както филмите са съставени от хиляди отделни статични кадри, някои хора са склонни да си представят, че времето е безкрайна поредица от отделни атоми и че движението се състои от миниатюрни стъпки, които могат да се възприемат само като непрекъснато движение. Този възглед е възприет за пръв път от философите от индийската религиозна секта Джайна в Северна Индия някъде около 200 г. след Хр. Според тяхната концепция един атом време е точка без размери, но същевременно той заема временен интервал, равен на минималната единица за време – точно както отделният филмов кадър не заема сам по себе си време, но успява да представи времеви интервал, когато бъде прожектиран.
Още преди появата на цивилизацията първобитните пастири са забелязали, че животните наследяват характерни белези от родителите си и че след няколко поколения желаните характеристики могат да бъдат затвърдени.
Изкуствена селекция
Ползвайки наученото, те са видоизменяли своите стада, като са отстранявали неподходящите животни и са получавали потомство само от избрани индивиди. Този еволюционен процес на изкуствена селекция ни е дал овцата (получена от дивата коза), кравата (произлязла от бизона) и прасето (произлязло от глигана).
Естествен подбор
Идеята, че подобен процес на естествена селекция е довел до развитието на добре приспособени видове, обитаващи днешния свят, е застъпена за пръв път в съчинението „Върху природата“ (V в. пр. Хр.) на гръцкия философ Емпедокъл от Акрага, автор на теорията за големия взрив. Неговата представа за еволюцията е твърде незряла – той предполага, че сред изчезналите, зле приспособими видове, изхвърлени при естествения подбор, може да е имало такива недоразумения като същества с глави, но без вратове или пък с ръце, но без рамене. По-късно гръцките мислители съставят значително по-добре обмислена теория. Естественият подбор потвърждава тяхната теза, че добре приспособените организми съществуват, защото сред случайно създадените при естествения градивен процес продукти те са успели най-добре да оцелеят и да се размножават. За поддръжниците на тази теория тя отрича вярването, че отделните същества са измислени и лично изработени от Бог. През I в. пр. Хр. естественият подбор отново намира своето място в „За природата на нещата“ от Лукреций:
„Преди един вид да оживее – той се нуждае от храна преди всичко и от части за възпроизводство,
чрез които семената на живота могат да намерят своя път
от мъжа към жената и да съединят телата
във взаимно удоволствие.
През онези стари дни
много опити бяха неуспешни, много от видовете
не успяха да оцелеят; това, което виждаме сега
да се радва на дъха на живота, трябва от първите
да е намерило защита за своя характер,
за своята сръчност, смелост и бързина.“
Идеите в това стихотворение са на гръцкия философ Епикур. Неговите научни схващания са заимствани най-вече директно от Демокрит и Левкип, създателите на теориите за атомите и молекулите. Вероятно един от тези двама гърци, живели през V в. пр. Хр., е първият, който е дал убедително обяснение за естествения подбор. Но тъй като техните трудове са изгубени, не можем да сме сигурни.
Произход на човека от маймуната
Големият ислямски учен Абу Рейхан Мухамед Ибн Ахмед ал-Бируни от Хорезм (973 – 1048) стига до заключението, че човекът е родствено свързан с други по-примитивни биологични видове. В своята книга „Китаб ал-джамахир“ той пише, че човечеството е резултат от еволюцията на дълга верига бозайници, най-близки от които са приматите (човекоподобните маймуни).
Еволюция на съвременния човек
Възгледът, че човешкият род е еволюирал от първобитния човек до днешния Homo sapiens, е бил изказан най-напред в Китай в сборника с научни и философски трактати „Хуай Нан Дзъ“ (II в. пр. Хр.), издаден в Нан Дзъ от Лю Ан по времето на династията Хан. Според тази книга човекът е минал през пет етапа в своята еволюция – започвайки от загадъчния „димен човек“ и достигайки до съвременния „обикновен човек“.
Дегенерация
Знанията за същността на еволюционния процес водят до появата на специфични възгледи относно бъдещето на човешката раса. Един от тях е политиката на евгеника, застъпвана от Платон. Според друг възглед многократното размножаване вътре в ограничен род води до дегенерация и непригодност. Тази идея се появява през 1813 г. в Англия в книгата „Лекции по физиология, зоология и естествена история на човека“, в която се твърди, че европейските управляващи среди дегенерират и е необходима тяхната подмяна. Английският председател на Камарата на лордовете не одобрява книгата я и обявява за противоречаща на Светото писание. Нейният автор – хирургът Уилям Лоуренс – се отказва от нея, за да запази престижа си. Но по-късно той е награден със званието барон и получава поста на старши хирург на кралицата, а Чарлс Дарвин преповтаря много от неговите изводи за естествения подбор в своето съчинение „Произход на видовете“ (1859). Дарвин не отдава нужното на Лоурънс, но признава, че еволюционната теория, развита в „Произход на видовете“, е много стара: „Много далеч от истината би било да се твърди, че този принцип е открит в днешно време… Принципа на селекцията аз видях ясно записан в една древна китайска енциклопедия.“
Този сайт използва ‘бисквитки’ (cookies), за да ви предостави възможно най-добро потребителско изживяване. Можете да промените настройките си за бисквитки, или в противен случай приемаме, че сте съгласни с нашите условия за ползване.ПриемамПрочети повече
Правила на поверителност
Privacy Overview
This website uses cookies to improve your experience while you navigate through the website. Out of these cookies, the cookies that are categorized as necessary are stored on your browser as they are essential for the working of basic functionalities of the website. We also use third-party cookies that help us analyze and understand how you use this website. These cookies will be stored in your browser only with your consent. You also have the option to opt-out of these cookies. But opting out of some of these cookies may have an effect on your browsing experience.
Necessary cookies are absolutely essential for the website to function properly. This category only includes cookies that ensures basic functionalities and security features of the website. These cookies do not store any personal information.
Any cookies that may not be particularly necessary for the website to function and is used specifically to collect user personal data via analytics, ads, other embedded contents are termed as non-necessary cookies. It is mandatory to procure user consent prior to running these cookies on your website.


 Символ на свободните зидари
Символ на свободните зидари Символ на свободните зидари
Символ на свободните зидари