Автор: Бранимир Руменов Велинов
Преживяванията на сетивността често остават непонятни за словото. Опитвали ли сте се да опишете дадена миризма, да я обрисувате в пълната ѝ форма и детайл, да я разкажете на свой познат? Когато говорим за миризмите, нашата лексика и способността ни да даваме на нещата имена ни предават, използваме мътни и отвлечени понятия, асоциации от целия ни житейски опит, за да достигнем до описание, чрез което останалите да съпреживеят случващото се във вътрешния ни свят.
Причината се крие в човешката физиология. Поради слабите физиологични връзки между обонятелните рецепторни неврони и мозъчните центрове на речта, обонятелната информация остава вербално неосмислена. Обонятелният сигнал, за разлика от всички останали сетивни сигнали, постъпващи в мозъка, директно достига кората на крайния мозък без да премине през таламуса, част от мозъка, която служи като кръстопът и разпределително звено, обединяващо информация от отдалечени мозъчни структури.
По странен и примитивен механизъм нервните импулси, които възникват в обонятелните неврони, се предават по аксоните им до обонятелните луковици (bulbi olfactorii), две издадени напред мозъчни образувания, толкова причудливи, колкото и еволюционно древни (фиг.1).
Оттам обонятелната информация поема по пътищата на лимбичната система, група от примитивни мозъчни структури, за които се предполага, че освен с обонянието са свързани с емоциите и спомените. Нервните импулси достигат първичната обонятелна кора, където се превеждат на древния език на миризмите, за да намерят своя собствен смисъл, неописан с човешки думи и неразтълкуван. Този донякъде примитивен механизъм разказва историята на отминали епохи в зората на еволюцията, на време, когато мисленето, такова каквото го познаваме днес, все още не е съществувало, а мозъкът се е лутал в тъмнината на един непознат свят, с обонянието като единствена ръководна светлина. Тогава животинските организми са разчитали на друга форма на мислене, обонятелното мислене и следвайки химичната логика на това сетиво успешно са общували със заобикалящата ги среда, за да се приспособят и да оцелеят.
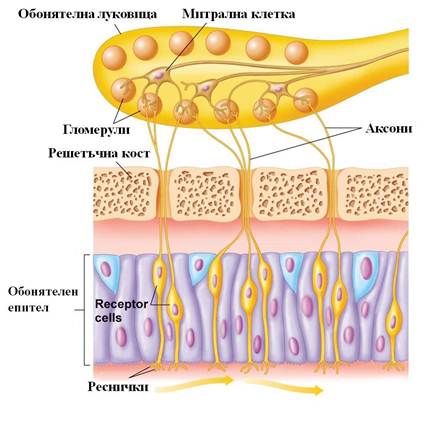
Фиг. 1 Организация на обонятелната система при човека. Обонятелните неврони (Receptor cells) взаимодействат с летливи вещества и предават сигналана обонятелните луковици в мозъка. (Principles of Human Physiology with Interactive Physiology, 4e (1))
Обонянието е част от инстинктивното ни и несъзнателно осмисляне на света, спомен от еволюционните уроци, които видът ни е усвоявал в продължение на хилядолетия. То разширява хоризонтите ни за познание: носи ни ценна информация за условията на средата и наличието на храна, алармира ни за опасност. Често обаче ни ограничава – живеем в затвора на собствените си възприятия. Спомнете си за половите феромони, доказани при редица безгръбначни и гръбначни животни, особено застъпени при насекомите. Езикът на половите феромони не оставя място за колебание – той заставя, ефективно предизвиквайки желания изход – намирането на полов партньор и успешното възпроизвеждане на вида. Използването на подобна химична комуникация със сигурност е ключовo за огромния еволюционния успех на насекомите.
А дали съществуват човешки полови феромони? Въпреки опитите на редица парфюмерийни компании да продават предполагаеми феромони, съвременната наука не е достигнала до изолирането и охарактеризирането им. Опити са правени – учените са си задавали въпроса за феромоните при хората и са достигали до разкриване на някои от тайните на човешкото обоняние. Линда Бък и Ричард Аксъл , носители на Нобеловата награда за медицина за 2004 г., правят едни от най-значимите открития за обонятелната система. Дали откриването на човешки полови феромони крие опасността да ни превърне в роби на собствената ни физиология, да ни лиши от право на избор, мога да запитам, илюстрирайки много характерна човешка черта – страха от неизвестното.
Много са загадките относно обонятелната рецепция, но неоспорима остава значимостта ѝ като огромно еволюционно предимство. Ако цветното зрението, друга важна еволюционна придобивка, може в редица случаи ефективно да определи годността на храната за консумация (зрелите плодове най-често са ярко оцветени) , то обонянието може твърдо да се произнесе по въпроса, различавайки класове от химични съединения (въглеводороди, алдехиди, карбоксилни киселини и много други). Миризмите не могат да бъдат описани с думи, но могат да бъдат категоризирани. Еволюцията на вида ни, индивидуалното ни развитие, както и рекламният бизнес дълго са отговаряли на въпроса кое мирише хубаво и кое лошо, поставяйки много от естествените миризми, които тялото ни излъчва, в категорията на неприятните.
Други аромати, например уханията на редица цветя са желани и търсени, предизвикват естетическо удоволствие, а често дори искаме да миришем на тях, т.е. ние се идентифицираме с тях. Като цветове на роза, ванилия или хелиотроп ухаят много от половите феромони на насекомите, предимно при пеперудите. Разсъждавайки върху този факт от еволюционна гледна точка, можем със същата увереност да кажем, че розите миришат на феромони на пеперуди: в продължение на хиляди години, цветовите ухания на растенията и феромоните на насекомите са коеволюирали в стремежа на висшите растения да привлекат опрашители. Парадоксално, миризмите, излъчвани от собствените ни тела ни отблъскват, но харесваме уханието на веществата, които насекомите използват, за да намерят полов партньор. Биохимичното и молекулярно познание относно обонянието крие тайните на непознати страни от същността ни, показва ни, че в природата абсолютните стойности са рядкост и всичко е относително, въпрос на отправна точка и индивидуално усещане.
Хеморецепцията, механизмът за детекция на химически вещества е възникнал рано в еволюцията, а степента в която е останал непроменен е изумителна: човекът, гръбначните и безгръбначните животни демонстрират изключително сходство в биохимичните основи на процеса. Обонятелните неврони при човека (фиг.2), разположени в горната част на лигавицата на носа, имат издължени клетъчни тела, завършващи с издължен израстък – аксон. В предния си край тези клетки имат множество реснички, подобно на морски главоноги, чийто пипала се извиват се и протягат се в океана на заобикалящата ни атмосфера, жадно търсещи контакт с подходящата химична молекула, която ще отключи усещането на дадена миризма.
В клетъчната мембрана на тези реснички са включени детекторите на всяка една миризма – молекулите на обонятелни рецепторни белтъци от семейството на G белтък-асоциираните рецептори (GPCR) . При човека са открити гени за повече от хиляда обонятелни рецептoра, като се смята, че само около 350 от тях са функционални. Линда Бък и Ричард Аксъл доказват, че всеки обонятелен неврон има в мембраната си само един вид обонятелни рецептори, а изходящата информация от всички неврони, съдържащи даден рецептор, се получава от една-единствена обонятелна гломерула в обонятелната луковица (фиг.1).
Нобеловата награда за физиология или медицина за 2004 г. е присъдена на Ричард Аксъл (Richard Axel) и Линда Бък (Linda B. Buck) “ за откриването на обонятелните рецептори и организацията на обонятелната система „.
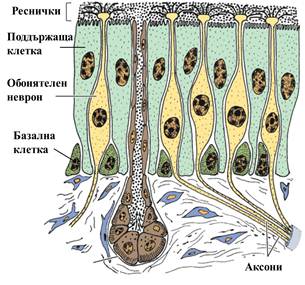
Фиг. 2 Обонятелната лигавица при човека. Обонятелните неврони имат издължени клетъчни тела с множество реснички в горния край. (Basic Histology: Text & Atlas,11e (2))
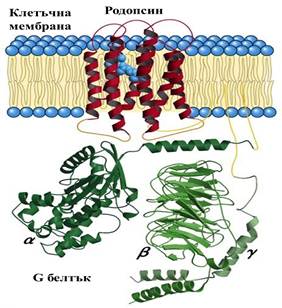
Фиг.3 Структура на родопсина и асоциирания с него G белтък. Родопсинът има характернаGPCR архитектура със седем трансмембранни α-спирали. (Lehninger Principles of Biochemistry, 5е (3))
Неизвестни остават точните триизмерни структури на обонятелните рецептори. Като съставна част на мембраната, те не са водоразтворими, което затруднява кристализирането им, необходимо за структурно изследване чрез рентгенографски анализ. Обонятелните рецепторни белтъци и останалите белтъци от семейството на G белтък-асоциираните рецептори имат изключително сходни аминокиселинни секвенции , а следователно и подобни триизмерни структури. Затова можем да визуализираме тяхната приблизителна молекулярна архитектура чрез разглеждане на изяснените архитектури на техни хомолози. На фиг.3 е представена структурата на родопсина от зрителната система, хомолог на обонятелните рецептори.
Полипептидната верига на G белтък-асоциираните рецептори, подобно на извиващото се тяло на змия, седемкратно пронизва мембранния бислой, откъдето идва и другото име за този вид рецептори – серпентинни рецептори. По този начин молекулата на рецепторния белтък образува дълбоко в мембраната канал за взаимодействие с други молекули (одоранти), всяка с ярко изявена химична индивидуалност, кодирана в определена структура, функционални групи и химичен афинитет. Всеки одорант представлява малка летлива органична молекула и участва в предизвикването на определено обонятелното възприятие. Механизмът на взаимодействие между обонятелните рецептори и одорантите е неизяснен. Методите за компютърно (хомоложно) моделиране се основават на структурното сходство между обонятелните рецептори и техните хомолози и имат известен успех в обясняването на свързването между обонятелен рецептор и одорант (фиг. 4).
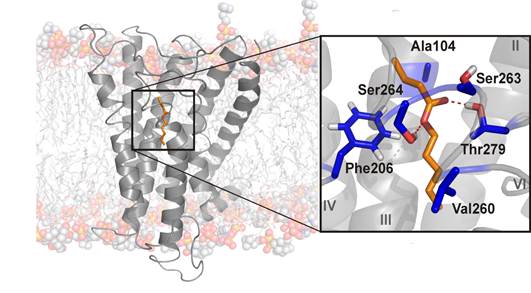
Фиг. 4 Компютърен модел на взаимодействие между обонятелен рецепторен белтък (сиво) и одорант (оранжево). Визуализирани са аминокиселинните остатъци (синьо) в свързващото място на молекулата на белтъка. (Prediction of a ligand-binding niche within a human olfactory receptor by combining site-directed mutagenesis with dynamic homology modeling, Gelis et al.(4))
Линда Бък и Ричард Аксъл демонстрират, че обонятелните рецептори показват групова специфичност по отношение на определен диапазон от одоранти: те доказват, че един одорант може да се свързва с различни обонятелни рецептори с различен афинитет. Според Бък и Аксъл различните миризми се кодират от различна комбинация от активирани рецепторни белтъци, т.е., обонятелната система използва комбинаторна кодираща система, за да различава обонятелните профили на веществата. Способността ни да различаваме миризми произтича от факта, че мозъкът интегрира входящата информация от различни видове обонятелни рецептори, за да разпознае „хибриден” профил, разширявайки диапазона на детекция отвъд броя на рецепторите. Интегрирането и комбинирането на информацията се извършва от гломерулите на обонятелните луковици, митралните клетки и други отдели на главния мозък (фиг.1).
Свързването на подходящ одорант предизвиква промяна на молекулната конформация на обонятелния рецептор (ОR) и задейства каскада от биохимични превръщания (фиг.5). Променил формата си, рецепторният белтък взаимодейства със следващия елемент от веригата – близко разположен G белтък , сходен по структура с G белтъка от зрителната система (фиг.3). Това взаимодействие, непонятно като механизъм за съвременната наука, предизвиква дисоциацията на α-субединицата на G белтъка и свързването на богатия на енергия гуанозин трифосфат (GTP). Освободена от белтъка, α-субединицата остава ограничена само от хидрофобна палмитоилна група, с която е „закотвена” към мембраната. α-Субединицата дифундира във вътреклетъчната среда, плъзгайки се по мембраната. Достига ензима аденилил циклаза (AC) и го активира. Активираната аденилил циклаза синтезира цикличен аденозин монофосфат (cAMP), вторичен посредник с разнообразни функции и огромно значение за живота на клетъчно ниво.
Аминокиселинната секвенция на един белтък представлява линейната последователност от аминокиселини, обуславяща първичната му структура.
Guanine nucleotide-binding protein – гуанин нуклеотид-свързващ белтък
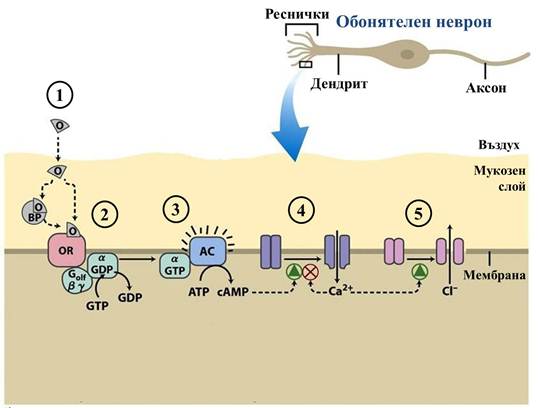
Фиг. 5 Молекулярен механизъм на обонянието. 1.) Одорантът (О) се свързва директно с обонятелния рецептор (ОR) или със свързващ белтък (BP), който го пренася до рецептора. 2.) Активираният OR предизвиква дисоциация на α-субединицата на G белтъка. 3.) α -Субединицата активира ензима аденилил циклаза (АC), който синтезира цикличен аденозин монофосфат (cAMP). 4.) Увеличената концентрация на cAMP предизвиква отваряне на катионни канали, през които навлизат калциеви йони (Ca2+) и деполяризират клетъчната мембрана. 5.) Калциевите йони отварят хлоридни канали. Навлезлите хлоридни йони (Cl-) деполяризират мембраната допълнително, задействайки изпращането на електрохимичен сигнал в мозъка. (Lehninger Principles of Biochemistry, 5е (3))
В мембраната на обонятелния неврон cAMP-регулирани катионни канали реагират на увеличената концентрация на cAMP, като се отварят. През тях навлизат калциеви йони (Ca2+). При покой, мембраната на обонятелния неврон е заредена, защото съдържа излишък на отрицателни заряди от вътрешната (цитозолната) страна. При навлизането на калциеви йони, носещи положителен заряд, потенциалната разлика от двете страни на мембраната се намалява, т.е. мембраната се деполяризира. Увеличената концентрация на калциеви йони действа като активатор на Ca2+-регулирани хлоридни (Cl-) анионни канали, през които хлоридни йони (носещи отрицателен заряд), напускат клетката и допълнително деполяризират мембраната ѝ. Деполяризацията на мембраната на неврона е електрохимичен сигнал, който достигнал критична стойност се предава на следващите неврони във веригата като потенциал на действие – от обонятелния неврон до кората на крайния мозък, предизвиквайки усещането на дадена миризма.
Познанието за обонятелната рецепция остава незавършено: неизвестни са структурите на обонятелните рецепторни белтъци, механизмите на взаимодействието между одорант и обонятелен рецептор, а съответно и механизмите за различаване на разнообразни миризми. В бъдещите изследвания е надеждата за по-голямо себепознание, за разкриване на молекулярната логика на важна част от същността ни, за разгадаване на езика на нямото сетиво.Библиография
- Stanfield, Cindy L. Principles of Human Physiology with Interactive Physiology 10-System Suite (4th Edition). Benjamin Cummings, 2011.
- Junqueira, Luiz C.; Carneiro, Jose. Basic Histology: Text & Atlas, 11th Edition, McGraw-Hill Medical, 2005.
- Lehninger, Albert; Nelson, David; Cox, Michael. Lehninger Principles of Biochemistry, Fifth Edition. New York : W. H. Freeman and Company, 2008.
- Gelis L, Wolf S, Hatt H, Neuhaus EM, Gerwert K. Prediction of a ligand-binding niche within a human olfactory receptor by combining site-directed mutagenesis with dynamic homology modelling, Angewandte Chemie, IE, doi: 10.1002/ange.201103980
- Ackerman, Diane. A Natural History of the Senses. New York : Random House, Inc., 1990.
- Buck L, Axel R. A novel multigene family may encode odorant receptors: a molecular basis for odor recognition. Cell. 1991, Vol. 65 (1), pp. 175–87.
- Buck, LB. The Molecular Architecture of Odor and Pheromone Sensing in Mammals. Cell.2000, Vol. 100, 6, pp. 611-618.
- Malnic B, Hirono J, Sato T, Buck LB. Combinatorial receptor codes for odors. Cell. 1999, Vol. 96(5), pp. 713-23.
Източник: nauka.bg
{kind=link}