 Филогеографията е нова дисциплина, обединяваща няколко дисциплини като генетика, молекулярна билогия, еволюционна биология, зоология, биогеграфия, ботаника и др. Стартираме с една преводна стария на човека дал името на тази дисциплина – Джон Авайз.
Филогеографията е нова дисциплина, обединяваща няколко дисциплини като генетика, молекулярна билогия, еволюционна биология, зоология, биогеграфия, ботаника и др. Стартираме с една преводна стария на човека дал името на тази дисциплина – Джон Авайз.
Джон Чарлз Авайз, роден през 1948 година е амеркански еволюционен генетик, еколог, консервационист и природен историк. Професор по Екология и еволюционна биология в Калифорнийския университет в Ървин. Той пръв дава името на новата дисциплина филогеография през 1987 година, като първите изследвания които прави в неговата лаборатория са посветени на генетичното родство и размножителното поведение при рибите и други животни.
Д-р. Чавдар Черников
/John C. Avise*.Phylogeography: retrospect and prospect. Journal of Biogeography (J. Biogeogr.) (2009) 36, 3–15/
Превел: Димитра Лефтерова
Въведение
Филогеографията е относително нова дисциплина, която се занимава с пространствения аранжимент на генетичните линии, по – специално между близкородствени видове. Името филогеография е дадено през 1987 (Avise et al., 1987). Приблизително десетилетие по – рано учените използват митохондриална ДНК за да изяснят как близкородствени видове са генеалогично свързани с вероятен общ предшественик/ митохондриалния геном е много подходящ за тази цел/. При генетични изследвания на различни видове са разкрити точни начини на пространствена подредба на мт ДНК генетични линии. Така да се каже, генеалогия се свързва с географията. След 1987 г., филогеографията става удобен способ за решаване на генеалогични въпроси на пространствените и времевите параметри на микроеволюцията. Във физиката Теорията на относителността на Алберт Анщайн ни дава концепцията, че времето и пространството са взаимосвързани и изкривяванията на времево – пространствения континуум могат предсказуемо да повлияват пътя на свободните частици. При филогеографията, няма подходящи математични закони, определящи точно начина на пространствената конфигурация на генетичните линии/ еволюционните процеси са твърде вариабилни и индивидуално различни за това/. Въпреки това за немалко видове сравнително филогеографски изследвания разкриват демографската и историческата природа на вътревидовата еволюция. Започвам този преглед с някой вече стандартни описания за изследвания на цитоплазмения геном и генногенеалогичната теория (Hudson, 1990), които най – често се използват при филогенетичните изследвания. След това ще илюстрирам, чрез някои практически примери ключови открития и новопоявили се принципи на филогеографията, преди да завърша с някои мисли за бъдещите насоки на тази процъфтяваща дисциплина. Тази статия предлага въвеждащи основи в областа, за повече и по – подробни обяснения и по – обширни цитати може да разгледате първата филогеографска литература Avise (2000).
Молекули и анализи
Митохондриалната ДНК еволюира бързо при животните
Митихондриалната ДНК ни връща обратно към протобактерия, която се е сляла ендосимбиотично с пре – еукариотна клетка рано в историята на живота на земята. По време на еволюцията, този цитоплазмен геном е загубил голяма част от своята ДНК и сега типично е представен в животинската клетка само с 37 гени всички свързани в една циклична молекула с приблизителна дължина от 17 000 нуклеотидни двойки. Два от тези гени кодират рибозомни РНК-и, 22 специфично различни транспортни РНК молекули и 13 кодират белтъчни субеденици които се колаборират с нуклеарно кодирани белтъци в биохимичните пътища на електронния транспорт и окислителното фосфорилиране, което помага за генериране на клетъчната енергия. Молекулата на мтДНК също така включва и контролен регион (CR) където се инциира репликацията на мт ДНК. Репликацията на мт ДНК е асинхронна с клетъчното делене и често в цитоплазмата на всяка соматична или клетки от гаметогенезата може да има стотици или хиляди копия.
Митохондриалния геном на животните е много компактен, не е обременен с интрони или дълги интрагенни участъци на некодиращи последователности, характерни за нуклеарния геном. Тези генетични облекчения трябва да са силно селектирани при конкурентните цитоплазмени условия, когато по – малките мт ДНК молекули вероятно се реплицират по – бързо, отколкото по – големите и следователно притежават значително предимство при предаването в поколенията.
Предвид важната функционална роля на мт ДНК в животинската клетка и генералната структурна икономика на клетката, преди 1980 г. е имало спекулации с това, че този геном би трябвало да еволюира бавно в нуклеотидната си последователност. Изненадващо, това не се доказва на практика: както първо показват Wes Brown et al. (1979), животинската мт ДНК акумулира нуклеотидни субституции неколкократно по – бързо отколкото едноверижна нуклеарна ДНК. Няколко фактора вероятно водят до бързата еволюция на мтДНК: относително недостатъчни механизми за поправка на ДНК в митохондриите; оксигенните условия в митохондриите обусловени от наличието на среда богата на кислород; липсата на функционални ограничения поради факта, че мтДНК кодира само няколко типа белтъци и не продуцира протеини директно свързани с нейната репликация, транскрипция и транслация; факта, че мтДНК е „гола” не е свързана с хистонови белтъци, които сами по себе си са консервативни и могат да ограничават еволюционното ниво на нуклеарната ДНК. Бързата еволюция на мтДНК е също така отразена в сегашно време сред много от животинските видове, с висока вариабилност на нуклеотидната последователност, основа на филогеографските анализи.
Популациите на мтДНК молекули присъстват в соматичните и пре – полови клетки
Големи популации на мтДНК молекули /често хиляди и повече/ типично присъстват във всяка клетка, така потенциално усложняват генетичните анализи, нещо което не съществува за нуклеарните гени. /Всеки автозомен ген, присъства само в две копия на всяка диплоидна клетка и в едно копие на всяка полова/хаплоидна//. Въпреки това на практика повечето от индивидите са хомоплазмени по мтДНК, което означава че техните мтДНК молекули са принципно идентични по нуклеотидна последователност. Хетероплазмени животни /тези които показват вътреиндивидуално различие на мтДНК/ са регистрирани рядко, и в тези случаи молекулите се различават по една или няколко нововъзникнали мутации.
Защо хомоплазмията предоминира е неясно, но е по – вероятна възможността мтДНК молекули при пре – половите клетки да преминават през относително популационно стеснение, такова че големия брой на молекули при всяка зигота да са получени от по – малко молекули в пре – мейотичните пре – полови клетки. Така всяка генетична транзиция от хетероплазмия в хомоплазмия или от хомоплазмия в хетероплазмия може да се получи от генетичен дрифт в относително малко на брой генерации животни/ както и се оказва експериментално/. Друга теоретична възможност е тази, естествения отбор да е в полза на хомоплазмията, или механизма на генната конверсия периодично да хомогенизира мтДНК последователности в клетъчните линии. Независимо от тези случаи, хомоплазмията е важна за филогенетичните изследвания, понеже всяко индивидуално животно носи един, специфичен митохондриален генотип.
Майчиното унаследяване характеризира животинската мтДНК
Друга отличителна характеристика на мтДНК е майчиното предаване на молекулата, тъй като цитоплазмата на зиготата основно произлиза от яйцеклетката. Всяко поколение получава мтДНК първоначално, освен ако не е изключение, от майка си, а не от бащата.
Женското предаване на мтДНК е подобно на мъжкото предаване на фамилните имена. В много човешки съобщества сина и дъщерята наследяват името на баща си, което само сина предава на поколенията. По същия начин синовете и дъщерите наследяват мтДНК от майките си, но само дъщерите ги предават на поколенията. Има няколко ключови различия: предаването на фамилните имена е условност приета от социума неотдавна /за пръв път се използва в Китай по време на Хан династията, приблизително в началото на новата ера/, докато предаването на мтДНК се случва от хиляди и милиони години; фамилните имена са характерни за хората, докато мтДНК „фамилни имена” присъстват природно при всички животински видове; освен това фамилните имена са съставени от няколко букви, докато мтДНК „фамилни имена” са съставени приблизително 17 000 нуклеотидни двойки /въпреки, че само част от тях се взема при типичните молекулярни изследвания/. Друга отличителна черта е, че всяко човешко фамилно име може да има няколко независими предшественици и така е полифилетично. Това се получава понеже източника на много фамилии е някаква професия /като Бейкър /пекар/ или Смит /ковач//, или видна историческа фигура / като Уилямс напр/, или превръщането на по – ранна система на патримония при която към името на даден човек се добява суфикс / Джонсон или Джаксон напр/. Обратно, всеки отделен мтДНК генотип принципно, с голяма сигурност е монофилетичен.
Понеже мтДНК се предава по единия родител, рядко, ако въобще някога рекомбинират. МтДНК се унаследява хаплоидно и безполово. Понеже при мтДНК няма пренареждането, което се случва по време на мейозата и сингамията, която се получава при сливането на половите клетки и засяга нуклеарната ДНК, мутациите които нармално определят генетичните вариации на мтДНК често се наблюдават в животинските популации. мтДНК генотипове се определят като хаплотипове, които варират помежду си с частични мутации натрупани и предадени от женския предшественик. Понеже мтДНК бързо еволюира, много и различни хаплотипове съществуват сред видовете.
Учените могат да използват тези хаплотипови последователности, за да определят майчината история на индивидите и популациите.
Генома на клетъчните органели има различни еволюционни правила при растенията.
Растенията имат два вида цитоплазмен геном – митохондриална ДНК /в митохондриите/ и хлоропастидна ДНК /в хлоропластите/ . Хлоропластите подобна на митохондриите също имат ендосимбионтен произход и също хлоропластидния геном е загубил част от гените си по време на еволюцията. Днес тези растителни органелни геноми са подобни на животинската мтДНК по своята структура /затворена циклична молекула/, начин на репликация /многобройни копия в клетката/, и не – Менделово унаследяване. Но също така те се различават от животинската мтДНК и помежду си по няколко важни молекулярни и еволюционни характеристики, както за пръв път е описал Palmer (1990).
Растителната мтДНК е много вариабилна по размер, варирайки от 20килобази до 2500 килобази при различните видове. Наследява се по майчин път, но не винаги така. Изненадващо растителната мтДНК еволюира бързо по отношение на генната подредба /генното реаранжиране е често срещано/, но по – бавно спрямо първичната нуклеотидна подредба. Така, еволюционната динамика на растителната и животниската мтДНК се различават диаметрално. Ниското ниво на еволюционна динамика на последователността на растителната мтДНК /около 100 по – бавна, отколкото на животинската/ плюс други технически усложнения означават, че тази мтДНК е слабо подходяща за филогеографски изследвания между видовете.
Растителната хлоропластна ДНК има други еволюционни правила. Тя слабо варира по размер между видовете /от 120 кб до 217 кб при фотосинтезиращите сухоземни растения/, като промяната на размера се дължи на удължаване на региона на дълга последователност от повтори. Молекулата има около 120 гени, които кодират рибозомни и транспортни РНК – и плюс някой полипептиди участващи в белтъчния синтез и фотосинтезата. Хлр(Хлоропластите)ДНК се предава по майчин път при повечето видове, от двамата родители при някой видове и по бащин път при други видове /повечето голосеменни/. Молекулата еволюира бавно, главно с генен реаранжимент, но също и с промяна на нуклеотидната последователност / около 3 – 4 пъти по – бързо отколкото растителната мтДНК, но много по – бавно от животинската/. Поради тези причини хлрДНК е доказано удобна за използване при оценка на филогенетични отношения при немалко таксони растения. С усърдно секвениране, достатъчно генетични вариации могат да бъдат разкрити, за филогеографски оценки между отделни видове.
Растителната хлрДНК принципно има много копия в клетъчната цитоплазма. Въпреки това повечето индивиди показват една хлрДНК хаплотипна последователност /и така са хомоплазмени/, а различните индивиди, между видовете често показват сериозни различия в последователността на хлрДНК. Така растителната хлр ДНК значително прилича на животинската мтДНК и учените могат по същия начин да използват нейния хаплоидна , нерекомбинираща природа, за да оценят генеалогичната история на растителните популации. Това проследяване може да бъде по майчина линия, по бащина линия или неспицифично и на двамата родители, в зависимост от начина на генетично предаване на хлр ДНК при отделните видове.
Нуклеарните локуси също имат генеалогична история
По принцип едно копие нуклеарни гени (scn) може също да осигури огромна последователност като основа за филогеографски изследвания на вътревидово ниво, но има три технически и биологически ограничения, които спират развитието на тези изследвания: относително ниското ниво на еволюция на последователностите на много от нуклеарните локуси; трудността да се изолират нуклеарни хаплотипове, от диплоиден организъм; феномена на вътрегенна рекомбинация.
Първото усложнение може да се превъзмогне с идентифициране на мониторингови scn последователности които еволюират много по – бързо спрямо останалите. Например интронните последователности на протеин кодиращите гени често еволюират по – бързо отколкото прилежащите им екзони. Втората пречка може да бъде преодоляна с използването на природната или изкуствена система на хаплотипната изолация. Например, гени на Х или Y- хромозомите на бозайниците /или Z и W – хромозоми на птиците/ са хаплоидни /хемизиготни в случая/ при всеки представител на хетерогаметен пол, както са всички scn локуси на мъжките от видове като пчелите или мравките с хаплодиплоидно полово определяне.. Когато гените в хаплоидно състояние не са осигурени природно /както са в случая на диплоидните организми/, е възможно да се пречисти нуклеарен хаплотип от всеки вид използвайки молекулярни методи /такива като изкуственото клониране в биологични вектори/. Независимо от използваните методи за генеалогични цели е важно да се изследват хаплотиповете индивидуално, защото иначе фазите на дадените хаплотипове /cis vs. Trans/ могат да останат неопределени при диплоидния индивид, което ще докаже хетерозиготността по много нуклеотидни позиции по продължение на гена.
Третата пречка заслужава уточнение защото недвусмислено е най –серозната. Във всяка популация на полово размножаващи се организми, мейозата и сливането на гаметите продължително пренарежда несвързани гени в нови мултилокусови комбинации. Тези рекомбинационни процеси могат да се случат и в рамките на даден локус, мокар че успешния мейотичен кросинговър се извършва между различни алели. Всеки рекомбиниран алел който се получава е като част на ДНК, омесена като амалгама, частите на която може да са имали напълно различно еволюционно минало. Така, вътрегенните /или вътреалелни/ рекомбинации се усложняват или представят неточно, което иначе казано е почистващ генеалогичен подпис от нуклеарния геном. Това контрастира с историческата чистота, характеризираща „ благословенно безбрачния” хаплотип на нерекомбиниращите цитоплазмени геноми (Dawkins, 1995). Въпреки това scn нуклеарни ДНК хаплотипове са използвани успешно при различни филогеографски изследвания (Hare, 2001), с едни от най – информативните резултати дошли от интрон последователности на протеин кодиращи гени. Понататък, дори и когато техническите и биологичните трудности на получаване на ясни генеалогични данни от scn нуклеарни ДНК последователности се окажат доказано непреодолими, не толкова директна информация за популационната история може все още да се добие от конвенционалните популационни генетични анализи, които се фокусират върху наблюдаването на честотата на филогенетични неподредени алели. Изследванията на алозими и микросателити, например, се използва рутинно за измерване на популационна алелна честота, което от своя страна може да бъде използвано при пространствено генетичните анализи на близкородствени популации. По специално когато такива данни са акумулирани на многобройни несвързани локуси, историческата информация често се случва да бъде сравнена с по – ясни генеалогични методи, които обикновенно се прилагат за цитоплазмените геноми.
Генеалогични концепции
Всички членове на популацията са свързани генеалогично
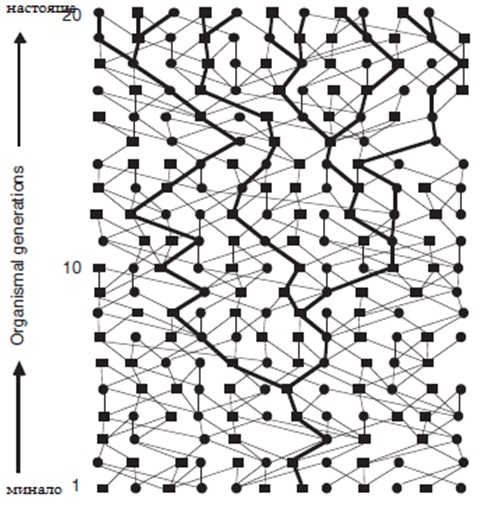
Въпреки многото различни генетични линии /аналогични на фамилните имена/, които могат да присъстват в популацията, те всички носят част от един общ предшественик , съществувал в някаква историческа точка. За да хванем тази концепция, разглеждаме първо майчината история на съществуващи индивиди при който е да е вид. Ако истинското популационно родословие е известно, всеки генеалог ще може да проследи майчините линии обратно във времето, и евентуално ще достигне до пра- пра – пра – пра – пра… пра бабата, няколко генерации назад, които са общи за всички сега съществуващи индивиди.Така всички сега съществуващи индивиди ще водят обратно в родословието до един общ предшественик по майчина линия. След това разглеждаме историята на съществуващата популация по бащина линия. Проследявайки обратно във времето по бащина линия и евентуално общия пра – пра – пра – пра…пра дядо ще бъде най – вероятно достигнат по подобен генеалогичен принцип. Важна особеност на майчината и бащина генеалогия, че те са като разклонено родословно дърво,/ не са мрежести/,въпреки че то е вградено в цялостното организмово родословие, което има твърде много разклонения вследствие от взаимното кръстосване. Всяка такава разклонена диаграма / събрани пътища, които са строго майчини или бащини/ , може да се счита като генно дърво или генетична генеалогия на организмовото родословие.
Трябва да се прави разлика между идеализираните генни генеалогии от една страна и не идеалните практически получени генни дървета от друга. Типична майчина генеалогия, например, е зададената по дефиниция действителна история на генетично предаване от майка на майка по генерациите, и типично бащино дърво е действителната история на генетично предаване от баща на баща на баща. Такива генеалогични истории не са познати директно от видовете в природата, но техните основни функции могат да бъдат оценени от подходяща ДНКпоследователност, сравнена между живи организми. Цитоплазмените геноми в частност могат да осигурят информация за майчината история, а Y хромозомата при бозайниците може да осигури информация за бащината история.
Клоните на генното дърво растат при всяка популация или видове
На пръв поглед може да изглежда така, че сумата от бащината и майчина история изчерпва всяка картина на генетично родословие. На това е далеч от истината на всяка популация на полово размножаващи се организми, защото повечето от нуклеарните гени са предадени и от двата пола. Представете си например факта ,че един индивид от неблизкородствено родословие има осем пра – пра родители и от всеки наследява приблзително 1/8 от неговата автозомална ДНК, така само два от тези пра – пра родители / 25%/ ще бъдат индивидуални майчини и бащини прародители.
Принципно всяко организмово родословие може да бъде декомпозирано на многобройни генни генеалогии / фиг.1/ които са аналогни на майчиното и бащино гореописано родословно дърво, но което ще включва предаването по бащин /М/ и майчин /F/ предшественик /както е за всеки нуклеарен автозомален локус/. Например един хипотетичен път може да включва редуващи се полове / F, M,F,M,F,M,F,M…/ от предходните поколения, а друг може да бъде M, M, F, F, M, M, F, F…. При всяко родословие което е G генерации дълго, теоретичния брой на отделни „ генни дървета” от този вид /както е определено от полово – специфични генеалогии/ е просто 2G. В реалния свят няма автозомен ген, който да е предаден само по майчина линия или само по бащина линия, вместо това и двата пола са включени /макар и не точно определен полово специфичен начин по няколко поколения/ . При всяко събитие, на всяко поколение, броя на предаваните пътища възможни за алелите на отделен автозомален ген е увеличен четирикратно (M fi F, F fi M, M fi M и F fi F) спрямо мтДНК(F fi F само) или Y хромозомата(M fi M само). Това също предполага че времето на предаване по генерациите на автозомален ген е очаквано четирикратно по – назад в историята , всички други условия са еднакви, от тези на мтДНК и на Y хромозомата. Тази базова читрикратна разлика също може да също да се види като произтичаща от двукратния ефект получен от диплоидноста на автозомалните гени / за разлика от хаплоидната мтДНК/ съчетан с друг двукратен ефект, резултат от предаването от двата пола /за разлика от предаването по единия пол на мтДНК и Y – хромозомата/. По – общо казано , многочислени отделни нуклеарни генни генеалогии съществуват в родословието на всяка полово размножаваща се популация. С други думи несвързани сегменти на ДНК имат квази – независимо историческо предаване, така както техните генеалогични структури се унаследяват различно, най – малко до известно родословие, от локус на локус.
Фиг. 1 Хипотетично генно дърво или хаплотипно дърво /по – платните линии/ на 20 поколения организмово родословие. Квадратите са мъжките, кръгчетата – женските, а линиите свързват родителите с техните деца. Концепцията на генните дървета и генеалогичните процеси са основни за появата на филогеографията като отделна дисциплина на биогеографията.
Генеалогията и историческата демография са преплетени.
Родословното сортиране съпътства организмовата репродукция и прави този вид генни генеалогии,обсъждани по – горе. По отношение на майчината генеалогия, например, родословното сортиране няма да може да се осъществи само ако всяка женска във всяка генерация се унаследи само от една дъщеря, при този случай няма да има йерахични майчини разклонения и предаване на наследствеността. Но в една реална популация, различни женски водят неравномерно до общия обем наследници, така че всяко майчино родословно дърво неизбежно се разклонява на няколко клони, наследниците се размножават или умират. Различните начини на размножаване поддържат неизбежния процес на предаване на поколенията.
За да илюстрираме това, да си представим че една женска роди дъщеря, съгласно статистически приетото разпределение на Поасон се означава /с отклоненията/ равно на 1.0. Поасон допуска че успешните дъщери, наследили майка си е независимо и случайно събитие. Съгласно този модел, очакванията майката да няма дъщери наследници в следващото поколение / или иначе казано очакването на майка без дъщери / е e=0.368 / където e е база на естествения логаритъм/ и нейната възможност за 1, 2, 3, 4 и 5 и повече дъщери е съответно 0.368, 0.184, 0.061, 0.015 и 0.004. Тези значения се добавят към едно поколение, но изчисленията / които са по – сложни/ се отнасят за много поколения. Те показват, например, че около 98% от всички основаващи майчини линии/матрилинии/ е вероятно да изчезнат след 100 поколения, дори и когато размера на популацията остава стабилен, а останалите 2% от матрилиниите оцеляват и продължават да се разможават.
Голям извод от такава статистическа теория е този, че исторически демографски фактори /такива като броя и вариациите на потомството на едно семейство/ може да се отрази на генната генеалогия по статистически придвидим начин. Например, точките на общите предшественици в генната генеалогия са по – стари в популациите с голям ефективен размер, при условие че всички други условия са равни. Така, практическите данни за дълбочината и разклоненията в структурата на генното дърво /като данни от мтДНК/ могат да се тълкуват съгласно генногенеалогичната теория , за да се изясни демографската история на популацията. Някой от прозренията на тази теория може да изглеждат нелогични. Например произхода от един предшественик в майчината генеалогия не предполага, че само една женска е живяла в това поколение, а просто показва че другите женски в това предшестващо поколение не са оцеляли до днешен ден с женски наследници. Друга важна точка е тази, че наследствието при майчината генеалогия не предполага, че другите женски в популацията са се провалили в предаването на генетичен материал на поколенията. Нуклеарните гени, от много от тези други индивиди могат добре да се представят в наследниците от алелите, които се предават не по майчин път в родословието. Аналогични твърдения и концепции могат да се направят и за бащините и всички други генеалогични пътища в рамките на разширяващото се животинско родословие.
Пространствено структурираните популации са генеалогично свързани
Филогеографските изследвания нормално се отнасят за многобройни популации разпространени по земята. Поведенчески и физически пречки на разпространение често са присъствали или присъстват в тези популации. Специално при тези видове с ограничена подвижност /възможност за придвижване/, популациите могат да бъдат частично изолирани една от друга на разтояние. Физическите филтри или бариери /такива като реки за сухеземните видове, или планини за равнинните видове/ често усилват ефекта на ограничената подвижност в развитието на популационната структура. Някой такива филтри могат да бъда полупропускливи и да позволяват случаен обмен на генетични линии, но други могат да блокират генния обмен въобще за дълъг период от еволюционно време. В последния случай, точката на общия предшественик за всяка генна генеалогия ще бъде не по – малко стара от появата на тази бариера. Целите на филогеографските оценки е да се използва генното дърво, за да се покажат историческите и съвременни сили, които образуват сегашната генеалогична архиктектура на популациите и близкородствените видове.
За всеки вид който генетично се разделя от някаква преграда на разпространението му, генеалогичния начин зависи от продължителността на действие на тази бариера, както и от историческата демография на разделените популации. Да разгледаме например , случайно чифтосващи се видове от предшествена популация, които се разделят на две отделни популации (A и B) от някакво изолиращо събитие. Първоначално е най – вероятно популациа А и В да са полифилетични спрямо майчиния произход /или мтДНК отношения/, което означава, че някой индивиди от А са генеалогично по – близко до някой индивиди от В, отколкото с други индивиди от А /или обратно/. След няколко поколения, при сортирането на отделните линии, в крайна сметка може да се получи така, че популация А да стане парафилетична спрямо В /или обратно/, което означава, че някой индивиди от А ще са генеалогично по – близко до някой индивиди от В, спрямо други индивиди от А, но всички индивиди от В са станали монофилетична единица по време на комбинирана генеалогия на популациите А и В. След още няколко поколения, А и В евентуално могат да станат реципрочно монофилетични по отношение на майчиния предшественик, което означава всички индивиди от съответната популация са едни на друг близки и генеалогично родствени по този специфичен компонент от родословието. Тази генеалогична транзиция от полифилия през парафилия до реципрочна монофилия в генното дърво е очаквана от генногенеалогичната теория, всеки път когато смесени предшествени популации се разделят на изолирани еденици /фазата на полифилия може да се пропусне, ако предшествената популация вече е била разделена преди това от някаква бариера/. Времева продължителност на транцизионните генеалогични етапи също зависят от големината на дъщерната популация, защото целия генеалогичен процес се повлиява от сортировката на линиите или реорганизацията на линиите произтича от диференциалната репродукция. Важна концепция е тази, че даден генеалогичен начин сам по себе си може да бъде еволюционно динамичен /по – скоро от статичен/ последван от събитие на разделяне на популацията.
Генеалогичните съответствия са доказателство за дълбочината на филогеографската структура
Предвид очакваната хетерогенност но генеалогичните пътища между видовете и многообразието на поведенчески, екологични и еволюционни сили които оформят генното дърво, как може филогеографските истории на популациите да бъдат реконструирани посредством молекулярни данни с голяма достоверност? Специфично, как може исторически дълбоки /древни/ популационно генетични подразделения да бъдат емпирично отличени от тези които са еволюционно плитки /скорошни/? Отговорите на тези въпроси се намират в принципите на генеалогичното съответствие (Avise & Ball, 1990).
По принцип, най – малко четири различни аспекта на генеалогичното съответствие или съгласуването сред белезите може да се определят.
Аспект I съответствие е в генния локус: ако много нуклеотидни субституции на последователността на ДНК твърдо отличават един хаплотип от друг, дълбоко /противоположен на плитък/ историческо разделяне може сигурно да се добави в разклоненията на генното дърво при изследванията. Това е истина понеже акумулирането на нови мутации е бавен процес, затова значително време трябва да е минало преди даден нерекомбиниращ ДНК хаплотип да покаже големи разлики в последователностите. Ако генеалогичното разделяне е документирано само за едно генно дърво, разделянето в това отделно дърво е можело да стане и по други причини /такива като балансиращия отбор, който може да е в полза на продължително възпиране на крайно отличаващи се алели на даден локус/.
Аспект II съответствие е в мулти – локусов план: ако дълбоко разцепване на много генни дървета твърдо отличава същия набор от популации, еволюционните сили отговорни за това, трябва да са имали значително геномно влияние. Най – походящия кандидат за такъв широкобхватен ефект на локуси е продължителното спиране на генния поток, такъв който възниква от продължителни физически, поведенчески пречки на разпространение. Така всяко емпирично документиране на съответствие в Аспект II, твърдо предполага дългосрочно историческо разделяне на популационно ниво.
Аспект III съответствие е в мулти видов план: ако пространствено последователно генеалогично разцепване се показва сред няколко близко разпространени видове, отговорните за това еволюционни сили трябва да са били със широко разпространен ефект на ниво биотична система или екосистема. Накрая, аспект IV съответствие се отнася за всяко м съответствие между няколко линии на емпирични доказателства. Например, ако генното дърво се резцепва при отделни видове и географски е следвано от продължителни препятствия в разпространението , които се знаят или се предполагат от независимо доказателство / морфология, подвидова таксономия или физическа геология/ дълбоко филогеографско разцепване между видовете твърдо може да се предположи. Някой от генногенеалогичните концепции, разглеждани в този раздел, може да изглеждат езотерични. Въпреки това те всички са заимствани от живия свят с интерпретация на практическите филогеографски модели, както ще покажа в следващия раздел.
Практически филогеографски модели
Здрава генеалогична стурктура характеризира слабо разпространените видове
Първите основни филогеографски изследвания на мтДНК включват югоизточния джобен гофер, доста седентарен копаещ гризач (Avise et al., 1979). В генетично изследване на 87 животни, разпространени в Джорджия, Алабама и Флорида, са определени повече от 20 различни мтДНК хаплотипове. Някой от тях се различават само по един или два изследвани нуклиотидни позиции, докато други имат по – големи различия. Всички тези мтДНК хаплотипове са доказано пространствено локализирани и близко свързаните хаплотипове нормално са географски близки. В добавка, дълбоко генеалогично разцепване отличава всички гофери от източната от тези от западната част на видовия ареал. Плитките разцепвания на генното дърво са показателни за рутинна изолация на разстояние, докато дълбоките разцепвания /изчисления на 3% нуклеотидна разлика/ вероятно регистрират по – дълга история на генетично разделение – може би повече от милион години от своя общ женски предшественик. Така ранни практически изследвания маркират раждането на филогеографията като отличаваща се дисциплина.
През последните 30 години, подобни филогеографски оценки, базирани на мтДНК последователности са проведени буквално стотици на слабо подвижни животински видове, различни малки бозайници, влечуги, земноводни до безгръбначни като сухоземни охлюви и щурци. За толкова разнородни организми, филогеографските резултати са типични като тези по- горе описани: за всеки вид, близки, подобни или идентични мтДНК хаплотипове са географски локализирани и малък брой /типично от 1 до 6/ дълбоки генеалогични разцепвания, обикновенно отделят регионалните популации, които вероятно се диференцират в отделни рефугии (Avise, 2000). По дълбоките разцепвания обикновенно се свързват с известно или вероятно препятствие за разпространение, такива като Панамския провлак, който се образува преди около 3 млн години и разделя много тропични морски видове /риби, бодлокожи и др/ на Атлантическа и Тихоокеанска популация, които сега са доста различаващи се помежду си.
Сходни филогеографски оценки, базирани на хлр ДНК са направени на няколко растителни вида. Например, между 85 популации на обикновенния бук генетично са изследвани в рамките на ареала на разпростаренение на вида в континетална Европа. 11 ралзични хлр ДНК хаплотипа са отличени (Demesure et al., 1996). Някой от тези хаплотипове са различаващи се по една или две позиции, докато други са различаващи се по повече. Главните открития са, че един хлр ДНК хаплотип предоминира в по- голямата част от Европа, докато други близкородствени на него са близко географски локализирани и един крайно различаващ се набор от хаплотипове има в Крим. По – широко разпространения хаплотип е вероятно последователността на предшественика от западна Европа /от който другите билзкородствени хаплотипове са възникнали с по – скорошна мутация/, а отличаващите се хаплотипове от Крим вероятно показват наличието на втори плейстоценски рефугиум за този вид в източна Европа.
Подвижните създания също могат да бъдат генеалогично структурирани
Много организми имат голям потенциал на разпространение, като възрастни форми /например крилатите насекоми, или рибите на откритите води/, като ювенилни форми /например морските безгръбначни с дълго живееща пелагична ларва/, или като гамети /например растителните видове с полен, разпространяващ се от вятъра/ Дори и при тези подвижни видове, исторически бариери на разпространение могат понякога да бъдат непреодолими. Например Панамския провлак блокира вътреокеанския генен поток на почти всички морски видове след появата си преди 3 млн.г. Това е рефлектирало в съвремието при голям брой от морските риби, скариди, морски костенурки и други видове, на които популациите от Атлантика и от Тихия океан са различни от двете страни на провлака. Те показват доста дълбоко историческо разделение при молекулярни изследвания на мтДНК и други гени (Knowlton et al., 1993; Bermingham et al., 1997).
Понякога дълбоко генетично разцепване разделя съществуващи популации, които сега не показват да са разделени от някакви пречки в разпространението. Типичен пример е
морската лещанка (Fundulus heteroclitus), която обитава амерканското атлантическо крайбрежие, където е регистрирано голямо генеалогично различие между северната и южната популация на основа на мтДНК и секвениране на нуклеарни гени, кодиращи лактат дехидрогеназа (Bernardi et al., 1993). Този пример на аспект II съответствие може да е доказателство на присъствие на видовете в два отделни изолирани рефугиума по време на плейстоцена където генетичната диференциация се натрупва, последвано от разширяване на ареала и вторичен контакт. Не може да не се каже, че естествения отбор също не оказва влияние за получаване на този краен филогеографски резултат. Например, естествения отбор в миналото може да развие някой от генетичните различия, които да останат в алопатрията, и обратно естествения отбор /от факторите на околната среда като температура на водата/ могат да инхибират хаплотипния обмен между северния и южния региони. Подобен пример включващ аспект II на съответствие, и за който подобни обяснения могат да се направят се отнася за ракообразното Tigriopus californicus, разпространено покрай американското тихоокеанско крайбрежие (Burton & Lee, 1994).
Някой организми с голям потенциал на разпространение могат да покажат очевидна популационна генетична структура, резултат по скоро от филопатрия /поведенческа вярност към определени места/ отколкото от физически бариери сами по себе си. Няколко китоподобни видове, например, матрифокални /социално организирани около женските/, и имат очакван генетичен почерк, който е матрилинеална популационна генетична структура. При мтДНК изследвания на гърбатите китове (Megaptera novaeangliae), популациите на всеки от трите океана са доказано подразделени по майчини кланове, които вероятно възникват и се поддържат от женската привързаност към отделни миграционни пътища. (Baker et al., 1993). Тази привързаност вероятно се развива, още когато малкото следва майка си в пътя на миграциите и, от водите на високи към ниски географски ширини. Мигриращите птици са също така силно подвижни, но няколкото изследвани вида показват значителна филогеографска структура на основа на гнезденето /а също така понякога и по миграционните пътища и местата на зимуване /. При тъмногръдия брегобегач (Calidris alpina), например, пет мтДНК клона отговарят на регионалните популации на гнездене в Европа, източен Сибир, централен Сибир, Аляска и Канада (Wenink et al., 1996). Този вид на популационна генетична архиктектура при мигрантите на дълги дистанции е очевиден резултат на верността на поколенията към отделните миграционни пътища.
Не всички видове са филогенетично подразделени
Въпреки че доста видове показват значителна филогеогорафска структура, съществуват изключения, и те могат да бъдат доста информативни за ширината на филогеографските принципи. Един вид, който видимо няма очевидно филогеографско разделение е червенокрилия дрозд (Agelaius phoeniceus), един от най – популярните птици в Северна Америка. В генетични изследвания на 127 индивида от целия континент, са определени 34 различни мтДНК хаплотипа,един от тях е най – срещащия се /практически повсеместен/ , доказано в сърцевината на „ звездоборазуването” на генеалогията, към който всички останали хаплотипове /всеки сам по себе си рядък/ са независимо свързани (Ball et al., 1988). Такъв „звездообразуващ” тип е очакван почерк за видове, които са се разпространили в последно време от един географски източник; най – често срещания и най – повсеместнен хаплотип е вероятно този на предшественика, от който по – редките хаплотипове са възникнали чрез отделни мутации. Така генеалогичните данни от мтДНК предполагат че, следвайки отеглянето на ледниците / започнало преди 18 000 г./, червенокрилия дрозд разширява широко неговия ареал и размера на популацията си, като колонизира континента от първоначален Плейстоценски рефугиум.
Друг пример от този основен тип е показан от американската змиорка (Anguilla rostrata). Този вид е катадромен, което означава че индивидите прекарват по- голямата част от живота си в реки и езера, но евентуално мигрират към морето, за да хвърлят хайвера си и умират /за разлика от анадромните видове като сьомгата, която живее в морето, но мигрира в реките, за да хвърли хайвера си/. Миграциите за хвърляне на хайвера от емериканските змиорки в Саргасово море, в западната тропична среда на Атлантическия океан, където мигрантите от източна Северна Америка се събират за случайни чифтосвания – панмиктични. Получените ларви после евентуално поемат по своя път обратно в пресноводните хабитати. Дали този странен жизнен цикъл означава, че ювенилните змиорки от техния широкообхватен пресноводен ареал са част от един добре смесен генофонд? Филогеографските данни получени от мтДНК и нуклеарни гени са напълно в съответсвие с тази възможност. Хаплотиповете и алелите са повсеместно разпределени /и във близки честоти/ сред изследваната популация на змиорките от Мейн до Луизиана(Avise et al., 1986).
За всеки такъв вид, които има един, добре смесен генофонд, генногенеалогичната теория, може да се прилага към генеалогичните данни, за да се оцени дали вида има дълговременен ефективен популационен размер. Ефективния брой на индивидите (Ne) се отнася за размера на идеализирана популация, която има същите генетични свойства /същото време на общо развитие/ като това, наблюдавано в реалната популация. Обикновенно, дългосрочно или еволюционно Ne е доста по – малко от N /размера на преброеното/ по една или няколко следни причини: броя на чифтосващи се мъжки може да варира от броя на чифтосващи се женски; популациите в природата обикновенно варират по размер във времето, и могат да преминат през известни стеснения; някои индивиди могат да оставят повече потомци от други, създавайки здравословно разнообразие в семействата; могат да се случат селективни помитания при които позитивно отбрани мутационни помитания се случват в популациите или видовете, като така свиват предшествениците до по – малко нидивиди, отколкото може да има; или други фактори, такива като периодичните изчезвания или реколонизации, които могат да доведат видовете да покзват по – ниско Ne отколкото могат да се предвидят от наличното преброяване. Генногенеалогичния подход за изчисляване на еволюционното Ne, включва оценка по молекулярни данни, хистограма на еволюционното време за поделения предшественик на всички двойки изследвани индивиди, и последващо сравнение на „несъотвестващото разпределение“ на наследствените очаквания /според неутралната теория/ за хипотетични популации с различни ефективни размери. За змиорките, чифтово изчислено по мтДНК последователности, разклоняване се превръща на времето за развитие на всички поколения като се има предвид, че продължителност на всяко поколение е 10 години и че тяхната мт ДНК еволюира със стандартна стъпка /2% секвенционна разлика на млн години/. Резултантното несъчетаващо се разпределение доказва, че е близко по сила и вид на очакванията за общото развитие на поколенията за една популация с еволюционно ефективен размер Ne = 5,500 женски (Avise et al., 1988). Качествено подобни изводи са направени за няколко други вида, които показват генетично добро смесване. Типично /както при американските змиорки/, еволюционно ефективните популационни размери са в един порядък по – малки, отколкото реално преброените популационни размери, може би защото всеки вид е минал през популационни стеснявания, един или няколко пъти в своята еволюция и/или поради други демографски фактори /упоменати по – горе/, които могат да намалят дългосрочно ефективния популационен размер под нормалните нива на преброяването.
Историчните процеси също се отразяват на биотичните съобщества
Няколко регионални, за дадена природа, генетични анализи сравняват филогенетичните изводи сред много близко разпространени видове (Bermingham & Moritz, 1998). Забележителни, но последователни открития са тези, че видовете с подобни ареали или екологии, често са генеалогично структурирани по подобен начин.
Това предполага, че исторически биогеографски процеси са не само видово – идиосинкратни по техния ефект, а вместо това имат също така съгласувано оформяне на генетичните архиктектури на дадените регионални биоти.
Един пример на аспект III на генеалогично съответствие е осигурено от генетични изследвания на крайбрежна фауна на югоизточните САЩ, където популации от Мексиканския залив при всеки от няколко вида са силно генетично различаващи се от доста популации по Атлантическото крайбрежие (Avise, 1992). Дълбоки генетични разделения между Атлантическите и тези от Мексиканския залив популации не са универсални при всички изследвани видове /американската змиорка дава точен пример/, но по способ на подреждане предполагат, че значителна част от крайбрежната биота е била генетично повлияна по един общ съгласуван начин. Филогеографските фактори отговорни за този случай не са напълно изяснени, но работна хипотеза е тази, че предшествени популации в Мексиканския залив и в Атлантика са исторически разделени, може би по време на Плейстоцена, и са натрупали генетични различия, които днес се поддържат от екологични препятствия /такива като океанските течения или разлики в температурата на водата/ за междурегионален геннен поток. Съгласувани генеалогични способи, за които историческите обяснения са по-ясни, включват флората и фауната в Европа. Генетични изследвания осигуряват сигурни доказателства, че три Европейски региони не са били покрити от ледници по време на Плейстоцена – Иберийския, Апенинския и Балканския полуострови – и са били първоначални рефугиуми за много животински и растителни видове, които в към днешно време реколонизират континента (Hewitt, 2000; Weiss & Ferrand, 2006). Филогеографски отпечатъци са очевидни днес при пространствения аранжимент сред много видове от различни генеалогични клонове /често наричани вътревидови „филогрупи”/, които ни връщат обратно през различни колонизационни пътища към един или два от тези плейстоценски рефугиуми. В много случаи, пост плейстоценския район на разширяване също води до такива отличаващи се филогрупи във вторичен контакт по границите на групите, където може да има хибридизация. Тези гранични зони са концентрирани в специфични ареали /отбелязани в Алпите, централна Европа и Скандинавия/, може би защото ледниковите рефугиуми са общи, историите на постглациална експанзия и на физически пречки на разпространение също.
Историята понякога се повтаря
Друга илюстрация на всичките четири аспекта на генеалогичното съответствие включва рибите от югоизточните САЩ. За всеки от няколкото вида изследвани по мтДНК , а някои и по нуклеарните гени, популациите в западните реки /които се отвеждат в Мексиканския залив/ са генетично различни от популациите в повечето източни реки /отвеждащи се в Атлантическия океан/ и полуостров Флорида (Avise, 2000). Подобни гранични зони също съществуват там, където два форми се припокриват и хибридизират. Тези открития твърдо предполагат присъствието на два отделни рефугиума, в които генетичните различия са се натрупали и от които е настъпило последващо разпространение. Въпреки, че тези филогеографски начини са генерално съгласувани сред много таксони, дълбочината на генеалогичното разцепления се различава значително от вид на вид. Една възможност е просто, че молекулярните еволюционни степени се различават при тези таксони, но е по интригуваща хипотезата че, тези различни съвременни видове може да са били исторически повлияни по различно време. През 2-те млн год на плейстоцена, контитенталните ледници са се образували и изчезвали циклично и така са ограничавали и освобождавали отделните рефугиуми. Може би някои от съвременните популации риби в източните и от западните водоеми водят обратно към един и същ рефугиум, но по различно време от глациалните цикли. За това ще е необходимо в един от рефугиумите по време на глациалните цикли предшествена популация да е изчезнала и да е заменена по-късно от колонизатори от друга. Мрежестия резултат би бил съгласуван в пространствен аранжимент на генното дърво, но с различна времева дълбочина на техните разцепвания. Тази вероятност, въпреки че е силно спекулативна, е също подкрепена от различни типове от биогеографични доказателства: структура на съобществата. Няколко видове риби в югоизточните/Атлантически/ или западните /Мексикански залив/ реки, предполагат че популациите някога обитавали една или друга от тези ареали са изчезнали /или може би са дивергирали толкова много, че им е даден нов таксономичен статус – вид/. Ако е така, това ще помогне да се обясни не само общите фаунистични модели, споменати по – горе, но също така защо тези две биогеографични провинции са принципно съгласувани с две главни молекулярни филогрупи, съществуващи сред отделните видове.
Дори и митохондриалното генно дърво само по себе си може да даде ключови биологични прозрения
Въпреки, че филогеографските анализи най – добре се базират на нуклеарни и на митохондриални генни данни, мтДНК генеалогия сама по себе си може да е достатъчна за някой цели поради своята специална връзка с женската популационна демография (Avise, 1995). Да си представим, например, един вид при който женските са седентарни, а мъжките се придвижват на големи разстояния и се чифтосват на случаен принцип с женски от различни места. Матрилиниите при тези видове ще бъдат точно структурирани, а нуклеарната генеалогия ще бъде географски разпределена на случаен принцип. Въпреки това крещящо разногласие между мтДНК и нуклеарните гени дърво, матрилинеарната структура сама по себе си означава, че всяка географска локация е автономна от гледна точка на размножителния резултат и това от друга страна може да има сериозни последствия за управлението на популацията и нейната консервация. За да се аргументираме по – конкретно, да разгледаме зелената морска костенурка (Chelonia mydas). Тези костенурки се събират по различни места от световните тропични океани и излизат на брега, за да снасят яйца. Иначе прекарват по – голямата част от живота си в морето, хранейки се на места, които могат да бъдат отдалечени на стотици и хиляди километри от местата за полагане на яйцата и така извършват Херкулесови репродуктивни миграции. Генетичните изследвания показват, че групите за полагане на яйца се различават едни от други по мтДНК хаплотипове (Bowen & Avise, 1996), което означава, че женските имат привързаност към местата за полагане на яйца /защото ако нямат, мтДНК хаплотипове трябваше да са смесени в различните места за полагане на яйца/. Така, ако костенурките от всички места за полагане на яйца се чифтосват на случаен принцип и техните нуклеарни геноми са измесени като резултат от това случайно чифтосване /а практически те не са/, превързаността на женските към местата за полагане на яйца, документирано от мтДНК ще показва значителна степен на демографска автономност за всяко място. Този извод има консервационно значение: ако на някое от местата за полагане на яйца се унищожи от природата или от човешка дейност, установяването на ново място на това място за гнездене е малко вероятно и още повече за кратко време. /Естествено, изоставянето на някое от тези места също се случва, но затова видът има много места за полагане на яйца/. Възниква и друг интересен контекст на матрилинеарната история сама по себе си. Зелените костенурки ни дават такъв пример. Едно от главните места за полагане на яйца на този вид е остров Асенсион, един малък скалист вулканичен остров намиращ се посредата между Южна Америка и Африка. Всеки сезон, женските, които иначе се хранят край Бразилското крайбрежие се отправят в едно 4 400 километрова одисея за полагане на яйца до остров Асенсион и обратно. Защо предприемат такова пътешествие, което изисква невероятни физиологични усилия и навигационни умения, когато има близки места за полагане на яйца, намиращи се по крайбрежията на Южна Америка? Една интригуваща възможност предположена преди години от Архи Кар /първия биолог, изучавал тези костенурки/ е че тези костенурки са започнали да полагат яйца на този остров още когато този остров е прото – остров преди 80 млн год.. Тогава Южна Америка и Африка са били разделени от много тънка морска ивица. През изминаващите хилядолетия континентите бавно са се раздалечавали от тектоничните движения, местата за снасяне на яйца на женските постепенно са се отдалечавали при всеки миграционен цикъл със сантиметри на поколение. Тази хипотеза, ако е истина, трябва да бъде потвръдена от ненарушена матрилиния от поколения предшественици прези окло 80 млн год до сега, които са гнездяли всички на остров Асенсион, и също така предполага голямо различие на мтДНК последователности акумулирани между тези костенурки, снасящи яйца на Асенсион и другите, снасящи айца в Южна Америка или другаде. Хипотезата на Кар е подложена на практическо тестуване и резултатите са фалшифицирани (Bowen et al., 1989). мтДНК хаплотипове на женските костенурки от Асенсион се различават от тези полагащи яйца в Южна Америка, но степента на различие на последователностите е много малка, показваща че матрилиниите на костенурките, полагащи яйца сега на Асенсион ни връщат към предшественик колонизатор много по – ранен, преди около 100 000 години най – много.
Преспективи на филогеографията
През последните три десетилетия филогеографските преспективи /митохондриалната и на други гени дървовидна система от които тези преспективи възникват/ добавят много неортодоксални прозрения за еволюционните генетични процеси (Avise,2007). Таблица 1 кратко сумира 21 такива концептуални и емпирични приноса. Филогеографията като дисциплина се развива фантастично през последните 30 години и не показва признаци на забавяне. Напротив, аз виждам възможности за разширение по най – малко три припокриващи се фронта.
Таблица 1. Много неортодоксални преспективи на еволюцията са възникнали от молекулярни открития на мтДНК и чрез филогеографски преспективи, мотивирани от тези открития. За пояснение, вижте долния текст и също така Avise (2000, 2007).
- Популационна йерархия. Всички популации на мтДНК молекули присъстват в соматичните и полови клетъчни линии и така добавят едно изцяло ново ниво на популационно генетичната йерархия.
- Стеснения при половите клетъчни линии. Хомоплазмията типична за животинската мтДНК, предполага че при популационни стеснения в половоклетъчните линии вероятно ще се предадат митохондриално наследствено.
- Консервативна функция и бърза еволюция. Бързата еволюция на животинската мтДНК е предизвикателство за идеята, че функционалните ограничения задължително налагат ниска скорот и ограничават промените в последователностите на ДНК.
- Асексуално предаване. Цитоплазмените геноми при видовете с полово размножаване нормално показват клонално / еднородителско, нерекомбиниращо/ генетично унаследяване.
- Матрилинеарни истории. Бързото ниво на мтДНК еволюция, съчетано с майчино унаследяване на молекулата, означава че могат да се възстановят вътревидовите майчини истории.
- Индивидите като оперативни таксономични единици. Понеже всяко животно има специфичен мтДНК хаплотип, всеки индивид може да се разглежда като една „оперативна таксономична единица” за генеалогични оценки.
- Концепция за генното дърво. При организмите с полово размножаване, мтДНК осигурява прототип пример за неанастомозираща или не – ретикуларна, немрежеста генна генеалогия, или” генно дърво”.
- Разширена концепция за генното дърво. По принцип, мултигенерационно родословие на всеки вид, размножаващ се по полов път съдържа в нуклеарния геном, многобройни неизоморфни генни дървета.
- Генни дървета срещу видови дървета. Генните дървета за несвързани нуклеарни локуси могат и често се различават топологично от съставна видова филогения; несъответствията между видовите и генните дървета, дължащи се на идиосинкратично сортиране на линиите, могат също да характеризират съществуващите таксони които са се отделили в миналото.
- Генеалогични топологии. Във връзка с дадени генни дървета, филогенетичния статус на всяка двойка родствени видове е само по себе си еволюционно динамично, с общ времеви ход на събитията, следващо за популацията или вида разделение, което да е полифилия, парафилия или реципрочна монофилия.
- Генеалогично съответствие. Поради идиосинкратичната природа на индивидуалните генни дървета, дълбоки разцепвания на популационно или видово ниво могат да се разпознаят само във връзка с някой от различните аспекти на „генеалогичното съответствие” /наличие на един и същ белег при двата алела , сред локуса, сред вида, или по различни линии на доказателства/
- Генеалогия и демография. Сред някои видове, процесите на развитие описват връзките между генеалогия и популационна демография и показват , че генеалогия и демография за са внимателно преплетени концепции.
- Генеалогия и пол. Ключови демографски и поведенчески параметри често се различават между женските и мъжките по начина на отразяване на полово – специфичните генеалогични структури.
- Ефективен популационен размер. Емпирични генно – дървесни изследвания, обяснени чрез теорията за съвместното развитие, показват че повечето видове имат изненадващо малки еволюционно ефективни популационни размери.
- Вътревидови популации. Емпирични данни показват, че отделнити видове често са подразделени на ясно разграничени генеалогични единици /вътревидови филогрупи/.
- Брой филогрупи. Емпирично, общия брой на ясно разграничените вътревидови филогрупи на вид обикновенно са малко (c. 1–6).
- Пространствена ориентация.Вътревидовите филогрупи най –често алопатрични и обикновенно са пространствено ориентирани по начин, които видимо е с добър биогеографски замисъл.
- История на природата. Генеалогичните данни показват това, че допълнително към историческите биогеографски събития, поведение и историята на природата на видовете също се отразява на филогеографските модели.
- Таксономия и консервация. Филогеографските данни и концепции са много уместни за таксономични решения и също така за консервационната биология.
- Филогенетични доводи. По принцип, и противно на конвенционалния разум, езика и концепциите на филогенията /генеалогията/ са силно свързани на вътревидово ниво.
- Микроеволюцията, подобно на макроеволюцията има история. По принцип, вътревидовата еволюция рядко образува вид равновесни резултати /такива като тези получени при островния модел на популационна структура/ които да са математично обяснени, обект на традиционната популационна генетична теория; вместо това вътревидовите генетични архиктектури са наследени исторически /генеалогично/ и идиосинкратично по биографичен контекст. Полезрението на филогеографията ясно разпознава и побира тази фундаментална реалност.
Разширяване на емпиричните анализи на многобройни нуклеарни локуси
Много филогеографски анализи досега са били фокусирани първоначално на цитоплазмените геноми /поради няколко логически защитени причини, дискутирани по – рано/. Получената информация от тези относително лесно получени резултати бе голяма, но в търсенето на нови средства, един значителен напредък в тази област носят генеалогичните резултати от изследването на потенциално интересния, но трудно достижим нуклеарен геном. Преодоляването на техническите трудности /такива като бързо изолиране на нуклеарни хаплотипове от диплоидни индивиди/ и естествените пречки /по – специално как да се справим с вътрегенната рекомбинация/ може да е обезсърчително, но всеки изследовател, успешен в тези начинания, веднага би станал научен пионер. Целите са, да се възстанови генната генеалогия от многобройни несвързани нуклеарни локуси с идеята да се търси модел на вътре – локусово генеалогично съответсвие / или липсата му/, което ще носи сериозни интрепретации на историчните записи на всеки вид.
Разширяване на генногенеалогичната теория
Генно генеологичната теория е възникнала първоначално с ориентация към един локус, така съществува една голяма стая за адресиране на вътрелокусови вариации в генногенеалогичните резултати като функция на различни генетични фактори /такива като степента на свързаност между локусите/ и за миналото и бъдещето на популационно демографските параметри. Значителна мултилокусова генногенеалогична теория също ще трябва да се съгласува към много варианти на пространствени популационни архиктектури, които природата може потенциално да сътвори. Свързана с тези усилия ще е необходимостта към по нататъшно интегриране /свързване/на очакванията от генногенеалогичната теория и други аналитични методи (Excoffier, 2004) с емпиричните данни в по тясно свързани, интелигентни комплекси. Целта на комплексите трябва да бъде твърда „статистическа филогеография (Knowles & Maddison, 2002; Templeton,2004; Knowles, 2008), която да отразява реалистична интеграция на популационната генетика и филогенетичните методи.
Разширяване на филогеографските анализи към други таксони
Въпреки, че стотици от практическите изследвания във филогеографията са публикувани, само една малка част от биологичния свят е изследвана по отношение на филогеографски преспекиви. Така че бъдещите усилия трябва да разширят филогеографските оценки към много други таксони. Доста от тази работа би била мотивирана от желанието да се разберат идиосинкратичните генеалогични истории на отделните видове, което може да е от специален интерес поради таксономичните неточности, консервационни отношения, очарователни природни истории или от други биологични мотивации, които да са видово специфични. Главния изследователски фокус също така може да е поставен върху сравнителни филогеографски оценки на мулти видови регионални биоти. Основани на минал опит /такъв като с различни таксони в югоизточните САЩ и в Европа/, поделени филогеографски модели /съгласуваности/ рефлектиращи на видимите исторически събития може би ще бъдат доказани в характеризирането на много видове от много регионални фауни и флори. Наистина, колаборативни усилия са вече напът да разкрият филогеографски модели на няколко континента и при различни субконтинентални региони, като целите им са да реконструират главните насоки на скорошновремевите истории на изолиране и разпространение във всеки един ъгъл на света.
Мястото на филогеографията сред науките за биоразнообразие
Преди началото на „филогеографската революция” в края на 1970г и на 1980г, относително малко връзки са правени между биолозите изучаващи вътревидовите /микроеволюционни/ генетични процеси и тези анализиращи надвидовите /макроеволюционни/ генетични модели. Досега тази област беше на популационните генетици., които се занимават с промените в популационните алелни честоти, произлели чрез мутации, генетичен дрифт, генен поток, естествен отбор и полов отбор. В последно време с това се занимават филогенетиците и систематиците, които работят най – вече с еволюционните отношения на видове и надвидови таксони. Основни дисциплини с които работи популационния генетик са статистиката, математиката, екологията и популационната демография, и вероятно с малко от филогегорафските принципи. Работата на систематика е свързана с отделни таксономични групи в музея или на полето, но вероятно с малко от принципите на популационата генетика. Това положение на отношения е и жалко и иронично, защото популационната генетика и филогенетичната биология трябва да са част от времевия еволюционен континуум. Понеже филогеографията се отнася до изследвания на близкородствени организми и свързани видове, тя би трябвало да се подразбира като клон от популационната генетика по своя специален фокус върху генеалогията /не само върху алелните честоти/ както и се вижда от историческите генетични записи в отделни части на ДНК. Най – точно казано, вътревидовата генеалогия може да се интрепретира като филогения, разглеждана по отблизо. Понеже филогеографията си взаимодейства с генеалогията / микрофилогения/ тя също може да се разглежда като клон и на филогенетичната биология. Но се отличава от традиционната филогенетика по своя специален фокус върху популационната история и демография. Традиционно настроение сред систематиците е, че филогенията няма значение на вътревидово ниво, понеже генетичните линии са гъсто преплетени и анастомозиращи при едни размножаващи се по между си популации. Както видяхме, региони от свързани нуклеотиди/ мтДНК или хлрДНК са само два примера/ имат дървовидна история сред видовете и разклонените структури на тези генни генеалогии носят значителна аналогия / макар и често в различен времеви мащаб/ на филогенетичните дървета сумирайки историческите взаимоотношения сред репродуктивно изолирани видове и таксони. Освен това, систематиците рядко разглеждат и имат предвид историческата популационна демография при реконструирането на видовите филогении, а наистина историческите демографии на родствени организми имат важно и продължително влияние на начина на наследствените вериги, свързващи популациите и видовете по време на еволюционния процес. Филогеографията и генногенеалогичната теория биха били изяснителни отколкото генеалогията и историческата популационна демография са, взаимно свързани. Накрая, филогеографията работи с пространствените и времеви параметри на генеалогията и така може да бъде разглеждана като клон на биогеографията /може би нейния най – естествен дом/. Тя отново се различава от традиционната биогеография по своя специален фокус на родствените популации и на подрабната генеалогична информация. Тя също се различава от екогеографията/ важен поддял на биогеографията; Gaston et al., 2008/ по своя специален фокус на историчните причини, като допълнителни към селектиращите сили и другите екологични процеси, опериращи през повечето съвременни времеви рамки. Посредством своите иновативни генеалогични преспективи и своите значителни връзки с няколко различни области, филогеографията ще продължи да осигурява здрав емпиричен и концептуален мост между няколкото главни еволюционни дисциплини които имат ограничена комуникация помежду си. (Fig. 2).
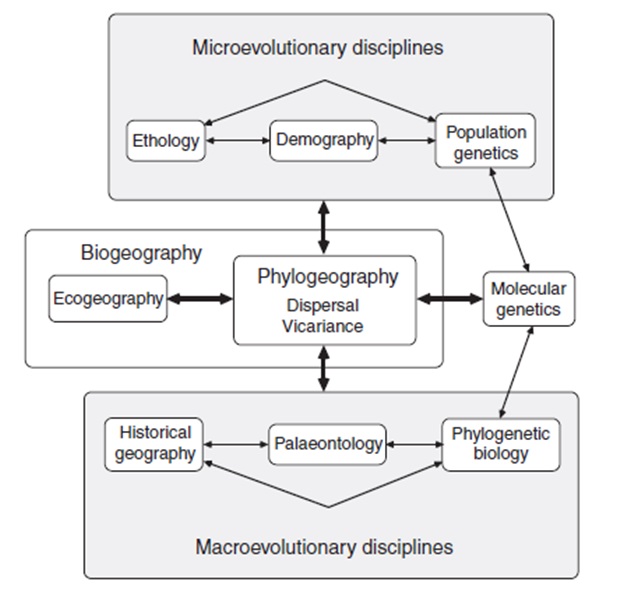
Фигура 2. Генералното място на филогеографията и някой от нейните емпирични и концептуални свързващи функции, сред науките за биоразнообразието (modified from Avise, 2000).
{kind=link}