Все повече днес сме залети от глупости, маскирани като наука. Телефонни линии на медиуми процъфтяват, политиците се съветват с астролози и хората отказват съветите на докторите си заради „алтернативно лечение“, практикувано от шарлатани. В миналото е имало защитници на истинската наука, които са разобличавали подобни нелепости.Къде са тези защитници днес?
За съжаление, те са заети да измислят собствени глупости.
Разбира се, това не е първия път в историята, когато хората вярват, че съдбите им са начертани на небето и болестите им могат да бъдат излекувани чрез молитви. Преди научната революция от 17-ти век такива идеи са били широко популярни. Жени са обвинявани във вещерство и са били изгаряни на клада. Прасета със странно поведение и кокошки с необичаен вид са били осъждани в демонично обсебване и са били екзекутирани.
После дошла Епохата на Разума, когато Исак Нютон призовал края на тези лудости. В най-известната си книга Principia той заявява, че „не измисля хипотези“ – т.е. не зачита никакви идеи, неподрекяпни от наблюдения. След Нютон, разпространителите на глупости са били изпратени директно в измерението на псевдо-науките.
До сега.
Днес, физиците смятат, че частица може да пътува по много различни траектории едновременно или пък назад във времето или внезапно да се появява или изчезва в нищото. Забавно им е да твърдят, че Вселената е просто „флуктуация на вакуума“ или като незначителен член на безкраен набор от Вселени или дори като холограма. Тъканта на тази странна Вселена се нарича „времепространство“, което се разширява, изкривява, посещава йога-курсове и може би има 26 измерения.
На кратко, последните публикации във физиката извикват носталгия за доста по-благоразумни неща, като съд на вещици.
В последното десетилетие все повече физици се разхождат по улиците със знаци „Краят на Физиката наближава“ в ръце. Твърдят, че са близо до разработването на „теорията на всичко“, която ще остави бъдещите им колеги с нищо за правене освен игране на видео игри. Можем да не обръщаме внимание на тази мегаломания, но пак бихме били изкушени да се съгласим с тяхното послание. Краят наистина изглежда близък, само че не в кулминационно издигане до пълно всезнание, ами срамно потъване в една псевдо-наука.
Този процес даже вече е започнал. Миналата година имаше много спорни дискусии около изследването на двама физици от Франция (братята Игор и Гричка Богданови). Проблемът беше тяхна публикувана работа, съдържаща спекулации за Вселената преди Големия Взрив, а спорът беше, дали работата е сериозна или пародия на съвременната космология. Истината се оказа по-страшна и от пародия: братята Богданови са съвсем сериозни, просто никой не успя да го види, така че колегите им бяха принудени да признаят, че много изследвания днес са неразличими от проста шега.
Физиците не са стигнали до това състояние за 1 нощ. Рано миналия век Айнщайн съвсем ясно отрече Нютоновия научен метод. „Сега осъзнаваме“, пише Айнщайн, „колко грешен е метода на тези теоретици, които съставят теориите си само от наблюдения.“ Вместо това той настоява, че теориите са „свободни творения на човешкия ум“. Неизбежният резултат от тази свобода е днешната „фантастична физика“.
Разбира се, физиците не признават, че участват в една фантастика. Те твърдят, че следват „хипотетико-дедуктивния метод“, което наистина звучи доста по-научно. Този метод им позволява да измислят каквато им допадне „теория“, стига да може от нея да бъде извлечено поне 1 последствие, което би могло да бъде наблюдавано някога, някъде от някой.
Истинското знание е трудно спечелена награда, която идва с постепенен процес от наблюдения през обобщения и съставяне на теория. За сметка на това, „измислянето“ на теории изисква малко усилие. Това обяснява, защо теоретичните физици успяха толкова бързо и лесно да достигнат „края на физиката“. За съжаление, техните измислени истории нямат отношение към хората в истинския свят.
Историята ни учи за важната роля на физиката в човешкия живот. В западния свят, познаването на физиката е издигнало човека от суеверен дивак, свит от страх пред природните феномени, до разумен мислител, който завоевава природата. Практическите ползи от тази трансформация са твърде много и твърде очевидни, за да се изброяват.
Но тук е заложено нещо повече от бъдещата технология. Докато наследството на Исак Нютон избледнява и физиката продължава отдалечаването си от реалността, нашата култура започва да губи от поглед най-ценната черта на човека: способността да мисли. Това е плашеща идея, защото ако човек не е „разумно животно“, то тогава той е просто „животно“.
Източник: nauka.bg
Автор: Дейвид Хариман (2003)
Превод: Мартин Сотиров
Учени за първи път свързаха електронно мозъците на двойки плъхове, давайки им възможност да комуникират директно, за да разрешават прости поведенчески пъзели.
В новопубликувано изследване, учени от университета Дюк излагат детайлно начинът, по който електронно са свързали мозъците на плъхове за първи път, позволявайки им дикектна комуникация при разрешаването на поведенчески пъзели.
Дърам, Северна Каролина – изследователи свързаха мозъците на двойки плъхове електронно за първи път, позволявайки им директна комуникация при разрешаването напрости поведенчески пъзели. При следващ тест тази връзка бе осъществена при две животни разделени на хиляди мили – едното в Дърам, Северна Каролина, а другото в Натал, Бразилия.
Резултатите от тези опити разкриват бъдещият потенциал за свързване на много умове, които да формират това, което изследователския екип нарича “органичен компютър”, и което може да позволи споделяне на информация от моторен и сензитивен характер между групи от животни. The study was published February 28, 2013, in the journal Scientific Reports.
“Наши предишни изследвания, включващи свързване на машина и мозък ни убедиха, че мозъкът на плъховете е много по-гъвкав отколкото сме предполагали.”, казва Мигел Никоелис, доктор на медицинските науки, главен автор на публикацията и професор по неврология в медицинския университет при Дюк. “В тези експерименти мозъкът на плъха се приспособяваше лесно, приемайки информация от външни на тялото му устройства и дори се научи да възприема невидима инфрачервена светлина, генерирана от изкуствен сензор. Въпросът който си зададохме беше: “Ако мозъкът е способен да възприема сигнали от изкуствени сензори, би ли било възможно също така да осъзнава информация подадена от сензорите на чуждо тяло?”
За да тестват тази хипотеза, учените първо тренирали двойки плъхове как да разрешат лесен проблем – да натиснат правилната ръчка, когато се включи индикаторната светлина над нея, награждавайки плъхът с глътка вода. След това свързали мозъците на две животни чрез внушителен брой микроелектроди, поставени в областта на кортекса, отговаряща за обработването на информацията свързана с двигателната дейност.
Един от двата гризача бил проектиран като “кодиращо” животно. То получавало визуално подсказване коя ръчка да натисне, за да получи като награда вода. Щом този плъх натиснел правилната ръчка, част от мозъчната му активност, кодираща поведенческите му решения бивала превърната в модел от електрически стимулации, доставени пряко в мозъка на втория плъх, познат като “декодиращо” животно.
Декодиращият плъх имал същите видове ръчки в неговата клетка, но не получил никаква визуална информация, за това коя да натисне, за да получи награда. Следователно натискането на правилната ръчка трябвало да се случи на базата на предадените сигнали от първия плъх посредством интерфейсът мозък-до-мозък.
След това, изследователите провели редица опити, за да установят колко добре се справя декодиращото животно в дешифрирането на мозъчния входящ сигнал от кодиращия плъх, за да избере правилната ръчка. Декодиращият плъх достигнал максимален успех от 70%, само малко под възможния максимум на успех от 78%, който изследователите предварително били изчислили на база на пряко изпращане на сигнали до мозъка на декодиращия плъх.
Важното е, че комуникацията осигурена от този интерфейс е двустранна. Например, кодиращият плъх не получавал цялостна награда, ако декодиращият плъх направел погрешен избор. Резултатът от тази особена ситуация, според Никоелис, довела до “поведенческо сътрудничество” между двойките плъхове.
“Ние видяхме, че когато декодиращият плъх правеше грешка, кодиращият буквално променяше мозъчните си функции, както и поведението си, за да улесни приемането на информацията от страна на партньора си”, казва Никоелис. “Кодиращото животно подобряваше съотношението на сигнал-звук на мозъчната активност, представляваща решението, така че сигналът ставаше по-чист и лесен за възприемане. Освен това правеше по-бързи и изчистени решения в изборът на правилната ръчка. Неизменно, когато кодиращият плъх правеше тези промени, декодиращият предприемаше правилното решение по-често, така че и двамата бяха награждавани.”
Във втората серия от експерименти, изследователите дресирали двойки плъхове да правят разлика между тесен или широк отвор, използвайки мустаците си. Ако отворът бил тесен, били научени да бутнат с нос канал за вода в лявата страна на клетката, за да получат награда, при широк отвор трябвало да бутнат канал в дясната страна.
Учените отново разделили плъховете на кодиращи и декодиращи. Декодиращите животни били научени да свързват определени стимулиращи импулси с правилен отговор отговарящ на левия канал за награди, а липсата на тези импулси с правилен отговор равен на десния канал за награди. В процеса на експериментите, в които кодиращият плъх установявал че отворът е широк и предавал изборът на декодиращото животно, то постигало успех в 65% от случаите, което доказва, че резултатът не е просто късмет.
За да проверят възможностите за пряката комуникация между мозъците, изследователите поставили кодиращ плъх в Бразилия, в Международния институт по неврология Едмънд и Лили Сафра в Натал (ELS-IINN), и предавали неговите мозъчни сигнали през интернет до декодиращ плъх в Дърам, Северна Каролин. В следствие на това, те установили, че двата плъха все още са способни да работят заедно по задачи, свързани с органите им за осезание.
“Въпреки че животните се намираха на различни континенти, въпреки шумът в сигнала и бавното му предаване, те все още можеха да комуникират”, казва Мигел Паи-Виейра, доктор, основен автор на изследването. “Това ни показва, че може би е възможно да създадем работеща мрежа от животински мозъци, разпръснати в различни локации.” “Тези експерименти представиха възможностите за установяване на изтънчена, директна комуникативна връзка между мозъците на плъхове, както и това, че декодиращият мозък работи като устройство, разчитащо образци. Така че, всъщност ние създаваме органичен компютър, който разрешава пъзели”, споделя Николелис.
“Но в този случай ние не вкарваме инструкции, а само сигнал, който представлява решение, направено от кодиращия, което се предава до декодиращия мозък, който трябва да разгадае загатката. По този начин, ние създаваме единична централна нервна система, съставена от мозъците на два плъха”, продължава той. Николелис отбелязва, че теоретично такава система не се ограничава до двойки от мозъци, а може да представлява “мрежа от мозъци”. Изследователите от Дюк и ELS-IINN сега правят експерименти да свържат множество животни, за да разрешават по-сложни поведенчески задачи.
“Ние не можем да предвидим какви качества ще се проявят, когато животните започнат да си взаимодействат като част от мозъчна мрежа. На теория, комбинацията от различни мозъци може да доведе до решения, до които индивидуалният мозък не би могъл да достигне”, казва Николелис. Подобна връзка може дори да означава, че едно животно ще бъде способно да инкорпорира в себе си чуждо усещане за “аз”, казва той. “Всъщност, изучаването на сензорния кортекс на декодиращото животно в тези експерименти показа, че в мозъкът на декодиращия плъх, в частта отговаряща за моторните функции, се представят не само личните мустаци на плъха, но и тези на кодиращият плъх. Засякохме неврони, които отговаряха и на двата вида мустаци, което означава, че плъхът е създал втора репрезентация на второ тяло над своето”. Основни изследвания на подобно приспособяване биха могли да доведат до ново поле, което Николелис нарича “неврофизиология на социално взаимодействие”.
Такива сложни експерименти биха били възможни в лабораторията, благодарение на способността да се записват мозъчни сигнали от почти 2,000 мозъчни клетки едновременно. Учените се надяват да запишат електрическата активност, произведена едновременно от 10 – 30,000 кортексови неврона в следващите пет години.
Подобен обеменен труд би направил възможен по-прецизния контрол върху двигателните невропротези – като тези, които се разработват от Walk Again Project (Проектът Ходи отново – бел.ред.) – за да възвърнат контрола на парализирани хора, казва Николелис.
Този проект наскоро получи субсидия на стойност 20 милиона долара от FINEP, бразилска агенция, която спонсорира изследователска дейност. Пари, с които ще бъде възможно развитието на първия цялостен екзоскелет, контролиран от мозъка, чиято цел е да възстанови подвижността на тежко парализирани пациенти. Първата демонстрация на тази технология е насрочена за откриването на Световното първенство по футбол през 2014 г. в Бразилия.
Освен споменатите Николелис и Паи-Виейра, други съавтори на изследването са Михаил Лебедев и Жинг Уанг от университета Дюк, както и Каролина Кюницки от ELS-IINN в Бразилия.
Изследването е било реализирано с подкрепата на Националния Здравен институт (R01DE011451), както и Националния институт за Умствено здраве (DP1MH099903), Фондация Вial, бразилската програма за националните институти по наука и технология, бразилския национален съвет за научно и технологично развитие и бразилските финансиращи агенции FINEP и FAPERN.
Publication: Miguel Pais-Vieira, et al., “A Brain-to-Brain Interface for Real-Time Sharing of Sensorimotor Information,” Scientific Reports 3, Article number: 1319 doi:10.1038/srep01319
Науката астрономия е започнала да се развива най-напред в Китай, а не на Запад. Това е изненадващо, защото в много други отношения старата китайска култура е била по-примитивна, отколкото водещите западни култури (в Месопотамия и Египет). Около 2700 г. пр. Хр. китайските астрономи са започнали да наблюдават и записват специфични явления на небето. Те са разделили небесната сфера на двайсет и осем сектора и са наблюдавали съзвездията във всеки от тях. Записвали са кулминациите на звезди във всеки сектор по време на равноденствие или слънцестоене. Определили са Полярната звезда и са следили регулярните движения на Слънцето, Луната и звездите. Според преданието, което няма как да бъде проверено, основополагащата работа е била извършена от „звездни служители“по време на управлението на владетеля Хуан Ди.
Нощна снимка на тамарау , Миндорски бивол (Bubalus mindorensis). Този бивол джудже е класифициран като застрашен от изчезване вид – което означава вид недалеч от етапа на изчезване от природата.
Камера поставена в планините на остров Миндоро от Филипините е заснела драматични снимки на тамарау, Миндорски бивол / Bubalus mindorensis – най-редкият бивол в света, ендемичен вид за остров Мандоро, Филипините, където е и най-големият вид на острова.
Таms-2 е амбициозен проект, участниците на който амбицирано искат да удвоят броя на тамарау от 300 на 600 до 2020г. Инфрачервените камери на Световният фонд за диви животни WWF, които са разположени навсякъде по света дават възможност за един поглед към тайният начин на живот на животните в природата.
Само 350 са останали живи
Камерите разположени на остров Миндоро показват откъслечни образи от тамарау – един труден и опасен за изследване вид. Само 350 от тези биволи джудета се смята, че са останали, съгласно Международният съюз за защита на природата и природните ресурси (International Union for Conservation of Nature, IUCN). Твърде опасно е този вид да бъде изследвам, с оглед безопасността на самият вид, застрашен от изчезване.
Някога през XIX век на острова е съществувала близо 10 000-на популация. След 1969 г популацията наброява под 100 индивида. (съгласно данни на WWF-Philippines).
За да се подкрепи запазването на вида и размножаването им в планинските хабитати , WWF с партньорството на Far Eastern University (FEU), (Отдела за околна среда и природни ресурси /DENR), Програмата за консервиране на тамарау (TCP), Hubbs-SeaWorld Research Institute (HSWRI/, местното управление на областта Миндоро, както и с участието на местното население в областта на националния парк Иглит Бако, е разработен амбициозният проект за удвояване на индивидите тамарау от 300 на 600 до 2020г.
Чумата по рогатият добитък, обезлесяването и лова
През XIX век на острова са ревяли близо 10000 тамарау. „През 1930г. е имало ужасяваща чума по рогатият добитък,освен това широко разпространеното обезлесяване и несекващото ловуване довеждат този вид до ръба на изчезването.” Обяснява Управителя на природен парк Иглит-Бако, г-н Родел Бойле. През 1969 г., популацията на тамарау наброява по-малко от 100 индивида, останали живи в горните тревисти части и малкото останали горски участъци на планините Иглит, Бако, Аруан и Калавит.
Благодарение на общите международни и местни усилия през април 2013 г. популацията вече наброява 345 индивида. „Тази инициатива се прилага не само за тамарау, но и за всички обитатели на парка”, добавя Бойле. Фантастичното биоразнообразие
„Миндоро е едно от седемте отделни биогеографски зони на Филипините. Западно Миндоро само по себе си е дом на две особено силно продуктивни природни зони – зоната на планинския парк Иглит Бако и Рифа Апо”., отбелязват заместник председателя на WWF Филипини и главния изпълнителен директор Хосе Ма и Лоренцо Тан. „Работейки съвместно с DENR / Отдела за околна среда и природни ресурси/, в нашият проект за съхранение на рифа, използвахме тамарау като пътеводен знак, за да възвърнем живота на планините, ерозиращи от оризовите полета в низините на Миндоро. Природно чистите реки, вливащи се в природно чисти крайбрежия, са изключително важни за поддържането на продуктивността на рифа, който от своя страна произвежда огромно количество морска храна.”
WWF, FEU и Отдела за околна среда и природни ресурси към Западно Миндоро интегрира консервационна програма свързана с тази за тамарау, като така подобрява управлението на парка и съдейства на инициативата за запазване на Апо рифа и богатството на морската флора и фауна в Саблаян. „Научно основаните действия водят до ефективно опазване.”, заключава Др Стюърт. „Тези снимки ни дават изключително важно виждане на движението и числеността на тези крайно потайни биволи. Когато ние знаем къде са, по-добре ще сме запознати кои райони да опазваме.”
Водорасли ни дават обещаваща възможност за борба с парниковите газове, съчетано с производство на биогориво
Вид водорасли ни показва възможност за редуциране на замърсяванията от парниковите газове, като същевременно произвеждат на биогориво, според ново изследване на Университета на Делауеър
Микроскопичното водорасло Heterosigma akashiwo бързо се развива в газова смес, която съдържа същите нива на въглероден диоксид и азотен оксид както емисиите изпускани от електроцентралите. „ Водораслите процъфтяват в газовата смес”, казва Катрин Койн, асоцииран преподавател по морски бионауки при Колежа по Земя, Океан и Околна среда към Университета в Делауеър. „Те растат два пъти по- бързо и клетките им са много по-големи сравнени с тези, които не са расли в такава смес.”
В Университа Делауеър изследователя Катрин Койн отглежда водорасли в лабораторията на университетският Люис кампус. Това може да помогне за намаляване на парниковите емисии, излъчвани от електроцентралите , като същевременно да бъде използвано за получаване на биогориво.
Водораслите също така произвеждат голямо количество въглехидрати, които могат да бъдат преработени в биоетанол като биогориво. Откритието може да има широко индустриално приложение като икономичен финансов начин за намаляване емисиите от парниковите газове, съчетано с производство на биогориво. Heterosigma akashiwo се среща навсякъде по света в природата. Койн, като експерт по цъфтежа на водораслите открива, че видовете имат специалното свойство да неутрализират азотния оксид – опасен за здравето на човека и околната среда газ.
Тази характеристика кара Койн и нейният екип да изследват дали водораслите могат да растат върху въглероден диоксид без да бъдат унищожени от високо съдържание на азотния оксид съдържащ се в газовите емисии на електроцентралите, както това се е случвало при опити на техни колеги с други видове водорасли. Близо година на лабораторни експерименти показва че Heterosigma akashiwo не само толерира газовите емисии, но и процъфтява сред тях. Водораслите не се нуждаят от допълнителен азотен източник освен азотния оксид за да се развиват, което значително намалява разходите за отглеждането им с цел получаване на биогориво. „Само това може да спести повече от 45% от необходимата енергия необходима за растежа на водораслите за биогориво”, казва Койн. Дотирани от Морската колежанска програма на Делауеър, Койн и нейният колега Дженифър Стюърт планират да продължат изследванията върху това как промяната на условията могат да увеличат растежа на Heterosigma akashiwo . Още повече, че те откриват сериозно увеличаване на нивото на въглехидрати, когато водораслите растат при наличието на парникови газове в сравнение с обикновен въздух. Те също виждат корелация между степента на осветление на водораслите и количеството на въглехидрати и липиди присъстващи в организмите. Изследователите разглеждат и възможноста за работа с компании за биогориво, за да могат да изяснят нивата на растежният процес и по- подробно да изследват Heterosigma akashiwo като вид за производство на биогориво.
Тази преспектива може да бъде подкрепена и в национален план от гледна точка на редуциране на нивата на парниковите газове, замърсяващи атмосферата, насока отбелязана в речта на Президента на САЩ Барак Обама, посветена на климатичните проблеми, която бе изнесена тази седмица. „Нашият подход към въпроса не е просто да се получава биогориво, а възможността да се използва този вид водорасло като биологичен пречиствател на парникови газове, с което да се намали тяхното вредно влияние.”, Казва Койн.
Видът Telekia speciosa е пример за вид, който не се е върнал в Западна Европа след последното заледяване, но който се разпространява от човека в градините. Тук е заснето във провинция в Дания. (Credit: Jens-Christian Svenning, Aarhus University)
Гората, която сме свикнали да виждаме не е изцяло в равновесие. След ледниковия период, някой растения са „изчезнали” в Северна Европа, т.е. видове които не са доживяли до наши дни. Същото се отнася и за други растения по света. Дори и днес има доказателства, че подобно събитие е възможно да се случи, но за много дълго време, под влияние на ледников климат или въобще климатични промени. В бъдеще такъв дисбаланс ще стане норма в растителните съобщества на Земята.
Двама изследователи от Аахус Университет – Дания, професора по биология Йенс Кристиян Свенинг и неговият помощник Броди Сандел, работят върху нова теория, анализирайки развитието на растителните съобщества от ледниковия период до сега.
Професор Свенинг обяснява „В дебата за климата, дори и при изследователите има тенденция да се омаловажава факта, че екологичната динамика може да е бавна. Нашите гори се адаптират изключително дълго време. Например, ние имаме все още малко на брой дребнолистни липи в Дания, които съществуват от периода на затопляне по време на Бронзовата ера, т.е около 3000 години. Може би, тя отново ще има шанс да се разпространи когато лятото стане по-топло. Но такава експанзия би била изключително бавна, тъй като тази липа не е бързорастящо дърво и не е бързо разпространяващ се вид, дори и при подходящи условия. Климатът ще се променя значително в рамките на живота на едно дърво, и ние не смятаме, че гората се е нагодила към новите условия. Бъдещият климат постоянно ще се изменя, което ще води до драстични промени и постоянен дисбаланс.
Дори и бързо разпространяващи се видове, като екзотични инвазивни видове растения, ще останат в дисбаланс за десетилетия и дори векове.
На шестила например, един високоинвазивен вид за Северна Америка са му необходими деситилетия, за да се разпространи дори на неголяма територия.
Предизвикателството пред нас
„Следователно, ако вие се опитвате да практикувате природен горски мениджмънт с природно възстановяване, може да видите възстановени обсолютно различни растителни видове в сравнение с тези които сте имали преди това, защото климата се променя така, че става подходящ за развитие на други видове. Това прави предизвикателство да се разработва и прилага в действие даден финансиран план за горски мениджмънт и регенерация. По същата логика, наличието на висок брой напълно развити застрашени растителни видове, не е гаранция че те ще присъстват в следващото поколение на съобществото. Това е предизвикателство за всички – за мениджърите, за хората които използват територията, за изследователите ,които работят с екосистемите, които са малко повече балансирани. Растенията и екосистемите като цяло, ще бъдат по-динамични и по несинхронизирани поради климата.
Ние сме предизвикали толкова много промени на климата, а същевременно природата е доста бавна в своето развитие. Помислете за продължителноста на живота на едно дърво. Нашата култура е основана на нещо, което не е в пълно равновесие, но е относително предвидимо. Свикнали сме със ситуацията където флора, фауна и климат са добре балансирани. В бъдеще, този баланс ще се промени въз основа на действуващите промени и ще настъпи сериозен дисбаланс. Ето защо ние се занимаваме с тази тема.”
Професор Свенинг също предупреждава: „В природата с такъв етап на дисбаланс, човешката намеса за интродукция на нови видове ще играе съществена роля. Например лавровишната, която може да се види в много градини в Дания, може да се разпространи в цялата страна. Но това разпространение на дивият тип, характерен за Юго Източна Европа към Дания, би отнело хилядолетия. Озеленителите доста помагат на процеса на разпространение. Това може да помогне на растенията да оцелеят, но също така може да предизвика по-бързо изчезване на северните видове от Дания. Лавровишната е вечнозелен храст и ако се разпространи , ще прави повече сянка на по -ниските видове в гората, което ще е от значение за тяхното оцеляване. В същото време такова разпространение би отнело десетилетия, в сравнение но това, способствано от озеленителите.“ Заключава професор Свенинг
Лавровишната е още един пример на вид, който не се е върнал отново в Западна Европа след периода на заледяване, но който ние разпространяваме в парковете и градините.
Спекулациите подобни на хипотезата за Гея, ни подтикват да не разглеждаха не традиционното разграничение между биологичното и физичното като нещо неразрушимо, особено на планетарно ниво. Ако физичните процеси, довели до възникването на Слънчевата система, съдържаха в себе си възможността да пораждат живот, както това е реализирано на Земята, то уместно е в изследването на физичната среда да се включи и изучаването на извънземните форми на живот. Знаем, че за да съществуват химични форми на живот, температурата не трябва да е много ниска, за да не замръзнат важните съединения, каквито са въглеродният диоксид и амонякът. Тя обаче не трябва да е и прекалено висока, за да не се разлагат прекалено бързо молекулите.
При много широко тълкуване на понятието за благоприятни условия този температурен интервал се простира между 200 К и 6000 К. Един по – реалистичен интервал би се оказал значително по – тесен, да речем от 250 К до 750 К. И в двата случая подходящите топлинни условия за живот в Слънчевата система съществуват само на малките вътрешни планети.
Ако една планета се държи като абсолютно черно тяло, тоест поглъща всичката падаща върху нея слънчева енергия и излъчва обратно в пространството топлината си под формата на инфрачервено лъчение, и ако разчита само на слънчевата радиация, тя може да достигне максималната стабилна температура. Ако разглеждаме тази планета като идеална проводяща сфера, която поглъща част от слънчевата енергия, а излъчва като абсолютно черно тяло, можем да изчислим нейната температура. За целта се използват данните за светимоста на Слънцето, обратнопропорционлната зависимост от квадрата на разстоянието и законът на Стефан. За планетите който имат атмосфера, температурата следва да се увеличи с множителя 2ј. Въз основа на такива изчисления се вижда, че само Меркурий, Венера, Земя и Марс предоставят условия за развитието на химчни форми на живот. От тези планети Меркурий изглежда напълно мъртъв, за този момент и Марс не изглежда по различно. Венера си остава загадъчна и негостоприемна и твърде възможно е живот да се е развил само на Земята. За момент ще оставим на страна хипотезите за юпитеровата луна Европа.
За да бъде наблюдаем животът в другите части на Галактиката, той би трябвало да е развил общество, чиято материална основа са науката и техниката. В противен случай как бихме могли да го забележим или да встъпим в контакт с него. Ако ние не считаме, че физичният произход на Слънчевата система е резултат от твърде редки и във висша степен неправдоподобни стечения на обстоятелства или пък, че произхода на живот на Земята е също така невероятен, то би трябвало да приемем, че Галактиката гъмжи от живот. Въпросът е до колко?
Разбира се, на този въпрос изобщо не може да се даде едностранен отговор. Неясните моменти са твърде многобройни. Най-многото което можем да направим, е да отделим факторите, които имат отношение по въпроса, биологичните и социологичните процеси, които са в основата на разбирането на проблема, и да преценяваме съдържателните догадки за горните и долните граници на техническите цивилизации. За момента има поне една такава цивилизация – нашата собствена цивилизация. Ако всяка звезда дава живот на една техническа цивилизация, то в Галактиката би трябвало да има 10ºº такива цивилизации. По такъв начин ние веднага можем да кажем, че броят на цивилизациите е между 1 и 10ºº. А може ли да се стесни този огромен интервал?
За образуването на една звезда са необходими десетки милиони години. Да предположим тогава, че при подходящи условия за около 1 000 000 000 се появява техническа цивилизация. Възрастта на Галактиката се изчислява на 1010 години тоест десет пъти повече от времето за създаването на една цивилизация. Ето защо честотата, с която се появяват техническите цивилизации трябва да е равна на честотата с която са се появили подходящи звезди преди около 1 000 000 000 години. Честотата на образуването на звездите, били те подходящи или не, е физична величина, която можем грубо да оценим по факта, че една звезда живее около 1010 и че в Галактиката има около 10ºº звезди.
Ако тази цифра характеризира едно равновесно състояние, при което умират толкова звезди колкото и се раждат, то среден живот от 1010 години на звезда означава, че годишно се раждат между една и десет звезди. Ако всяка звезда ражда техническа цивилизация, то горната величина би отразявала и честотата, с която възникват и техническите цивилизации. Съвършено ясно е, че е невероятно да се реализира тази ситуация в действителност. Не всички звезди формират планетни системи, а по-масивните пък могат да еволюират прекалено бързо, за да може животът да се развие в достатъчна степен. Дори и когато съществуват планети, по един или друг начин животът може да не достигне интелектуалното ниво, което е необходимо за наука. Разбира се, подобни фактори трябва да намалят честотата на възникване на технически цивилизации поне десет пъти. По такъв начин годишно могат да се появяват не повече от една напреднала цивилизация. Това е максималната честота. Оценките за тази величина, правени от астрофизиците (в това число Сейгън, Школовски) варира между 1 и 10ˉІ годишно. Истината е, че никой няма каквато и да е ясна представа за всичко това.
Какво може да се каже за средната продължителност на живот на една техническа цивилизация? От кога датира галактичната същност на нашето собствено общектво? От откриването на радиото? Във всеки случай става въпрос за възраст по скоро десетки от колкото стотици и хиляди години. Но това време е достатъчно, за да излязат на яве множество опасности, на които е изложено едно технологично общество – ядрена и биологична война, замърсяване, пренаселване, които могат да изтощят, и макар и не винаги, да унищожат човечеството. Продължителноста на живота на напредналите общества може би не надвишава няколко стотици години. Техни паметници може би са безплодните прашни планети. А може би пък животът на една такава цивилизация продължава докато еволюцията на нейното слънце и позволява, това е време от порядъка на 1010 години. За сега тук се крие най – голямата неопределеност. Каква е средната продължителност на живота на високо развитите технологични общества? Да се задава този въпрос на съвременната социология е все едно да се задава на Талес Милетски вапросът, какъв е периода на полуразпад на уран 238. Оценките за средна продължителност на цивилизованите общества проявяват склонност да варират в интервала между 10 000 и 10 000 000 години, а това грубо казано е и еволюционната времеконстанта. След 10 000 000 години обществото вероятно еволюира далеч от научнотехнологичната си фаза към нещо съществено различно. Вероятно е и подобно на динозаврите то да загине. Търсейки истината пипнешком, в своето невежество ние може би бихме могли да оценим и по – зле правдоподобната горна граница на продължителността на една цивилизация от тези 10 000 000 години, които възприемаме вместо първоначалните 1010 години. Ако сме крайни песимисти, ние можем да приемем, че долната граница е 10 000 години.
Като умножим честотата на възникване на технически цивилизации по средното време на техния живот, получаваме постоянния им брой в Галактиката. Максималния брой се получава при умножаване на максималната честота от една техническа цивилизация годишно по по максималната продължителност на живота им от 10 000 000 години. Това дава 10 000 000 цивилизации в Галактиката. Минималният брой се получава като вземем минималната честота от 10ˉІ цивилизации годишно и я умножим на минималното време на живот от 10 000 години. Това дава 100 цивилизации. Тогава след множество уговорки можем да твърдим, че по всяко време в Галактиката има между сто и десет милиона технически цивилизации.
Ние можем да използваме среднана плътност на звездите, за да оценим средното разстояние което разделя планетарните общества. Ако те са само сто в цялата Галактика, то всяка от тях е отдалечена от най-близкия си съсед на около 4000 светлинни години. Ако обаче те са 10 милиона, отдалечението е само 100 светлинни годин. Но дори и 100 светлинни години е значително разстояние. Ако електромагнитното лъчение е в действителност най – бързия способ за пренос на информация във физичният свят, то 200 годишната пропаст между въпрос и отговор не дава никаква възможност за остроумни диалози. В наши дни в отсъствието на измислените от фантастите хиперпространствени преходи и мигновени предавания на информация, дори и да съществува цивилизация, подобна на нашата, на разстояние стотици или хиляди светлинни години, ние не можем да я разглеждаме като нещо повече от обикновен интелектуален куриоз. Струва си обаче да се замислим за безкрайните възможности които процесът на еволюцията може да породи, тъй като тези възможности биха могли да се реализират някъде в нашата безкрайна галактика. Това възраждане на елементи на древногръцката философия и по специално на Платоновото твърдение, че всяка интелектуална форма на живот има свое материално въплащение, води до отрезвяващата мисъл, че може би дори ние все още сме на твърде примитивна фаза на развитие, за да можем да възприемаме природата и моща на високо развитите галактични култури. Ако всичко, което може да еволюира, в действителност еволюира някъде, то може да се окаже твърде различно от това, което ние си представяме или бихме могли да си представим. Така стигаме до още по – отрезвяващата мисъл, че то може да присъства някъде около нас и въпреки това ние бихме могли да не го разпознаем.
През 1801 г. Пиаци открива Церес – най-големия от астероидите, а през 1802 г. Олберс открива Палас и Веста – следващите два най-големи астероида. Този астроном известен с парадокса си, изказва хипотеза, че астероидите са останки от някогашна планета. Според хипотезата му, която и днес не е опровергана, орбитата на тази планета е лежала между орбитите на Марс и Юпитер. Произхода на астероидите и до днес остава загадка.
Церес
През 2005 г. списъка на астероидите с определени орбити наброяваше 214 000 тела по-голямата част от тях са скални късове с диаметър около 10 км. но диапазонът им варира от 380 км. за Церес и 240 км. за Палас и Веста до радиуси по малки от километър. Техните орбити повече или по-малко лежът в равнината на еклиптиката с наклон към еклиптиката точно 35°.
Като цяло поведението на повечето астероиди пасва с общата картина на Слънчевата система и те представляват много малка заплаха за навигацията. Всъщност именно поведението на няки от по-малките астероиди дава интересни аргументи в полза на поддръжниците на космическите технологии.
Bеста
Орбитата на Ерос не е много по-елиптична от тази на Палас но тя е много по-близка до тази на Земята. Средно тя е по-близка до Земята дори и от Марс. Така елиптичната орбита на Ерос го довежда приблизително веднъж на седем години на разстояние около 25 милиона километра. Това е безопасно разтояние до нещо което малко се отличава от извънземна скала с размери
7 км. х 19 км. х 30 км.
Ерос
Значително по-опасни са близките траектории на трите по-малки астероида радиусът на които е само няколко стотин метра. Те имат неправилна форма и непрекъснато се премятат докато профучават покрай Земята. През 1932 г. Аполо не ни улучи като отстоеше само на 3 милиона километра. През 1936 г. Адонис се доближи на 2 милиона километра а през 1937 г. Хермес постави рекорд които до 1977 г. не беше надминаван. Той достигна 750 000 км. около два пъти разстоянието до Луната. Адонис и Аполо и около десетина още техни компаньони редовно повтарят това свое тревожно поведение. Друга потенциална заплаха е астероида Икар – тяло с радиус около 700 метра, чиято орбита е изключително добре очертана. Средното разстояние между него и Слънцето е същото както и при Земята, от тази гледна точка Икар е изключително близък съсед. Орбитата му обаче е прекалено елиптична и в по-голямата част от времето той е на безопасно разстояние. Когато беше открит през 1949 г. той се намираше на 6 милиона километра. Орбитите на Земята и Икар не се пресичат точно но е нужна съвсем малка претурбация за да се осъществи нежелания контакт. Икар обикаля около Слънцето за 408 дни или 1.12 години, това означава, че на веки 19 години той и Земята силно се сближава. Две такива сближения вече имаше през 1968 г. и 1987 г. като първото някои хора ще помнят дълго.
През 1968 г. астрономите със затаен дъх проследявали приближението на космическия пратеник Икар. Заради грешка в изчисленията била и предвиждана точната дата на сблъсък със Земята – 14 юни. А размерът на подобна катастрофа би бил зловещ. Бихме ли могли да си представим размерите на катастрофата ако в Земята се забие тяло с маса 4,5 милиарда тона движещо се със скорост 155 000 км/ч би се образувал кратер с размери повече от сто километра в диаметър а взривът би унищожил всичко в огромна площ около него. Ако Икар или друг подобен обект се открие, че се приближава опасно близко има обсъждани възможности за кацане върху него и монтирането на мощен ракетен двигател с чиято помощ тялото да бъде отхвърлено на далеч. Следващото сближение с Икар ще е 2006 г. (тоест тази година) но за момента астрономите не засичат нищо тревожно.
Но не винаги Земята е избягвала срещи от подобен род. Една такава среща се е състояла на 30 юни 1908 г. На тази дата не голям астероид (или комета) с маса няколко десетки хиляди тона и радиус около 80 метра се врязва в земната атмосфера със скорост 30 – 40 км/сек. На височина около 6 км. от повърхността на Земята в района на река Подкаменная Тунгуска (Сибир) небесното тяло се взривява. При този взрив се отделя енергия равняваща се на 3 000 000 тона тротил или сто пъти по голяма енергия от отделената при избухването на атомните бомби над Хирошима и Нагасаки. В ройона на взрива дърветата в огромна площ от 20 000 кмІ били повалени в радиална посока.
Дълго време остатъци от това небесно тяло не били откривани, чак през 1980 г. участниците в експедицията на Института по геохимия и геофизика на Украинската академия на науките успяват да открият диамантено – графитни кристали с извънземен произход. В торфетия слой на катастрофата бил намерен и значително количество тежък въглерод С-14. Като се има в предвид разсейването на изотопите и мощноста на експлозията учените изчисляват теглото на Тунгуския меторит. По своите мащаби падането на
Тунгуския метеорит е една от най-големите катастрофи в съвременната история на Земята.
Българският принос – А79
От казаното по–горе недвусмислено става ясно, че изледването на малките тела в Слънчевата система е сред най – важните дялове на астрономията в които дейно участие могат да вземат и любителите астрономи.
През 2005 г. у нас се създаде и защити статута си нова обсерватория-Zvezdno оbshtestvo Obsevatory, Plana- MPC A79. Понастоящем тя е втората обсерватория след НАО ”Рожен” към IAU в България. Minor Planet Center (MPC) е центъра за малки тела в Сл. система и е основно подразделение на Между-народния астрономически съюз (IAU) – базиран в Харвард, САЩ. Това е организацията, която е оторизирана в изследването на движението на малките тела -астероиди и комети. По време на наскоро провелия се конгрес на IAU в Прага, се дефинираха и нови класове от обекти като например планети-джуджета (Плутон, Цереа и др.), което предизвика бурни дискусии както в астрономическите среди.
А79 е създадена и ръководена в момента от астронома Филип Фратев, който има дългогодишен опит в тази област на астрономията и е участвал в наблюдателният екип на ”група малки тела в Сл.Система” към института по астрономия, БАН в периода 1990-1994 на обсерваториите Рожен и Белоградчик. През 2004 година създава малка обсерватория към MPC- МPC A61 (Tortona) която се намира на около 60км от гр.Милано, Италия.
Както в Италия (MPC A61) така и тук (MPC A79), дейността на обсерваторията е финансирана на собствени начела и е свързана предимно с изследване на движението и определяне на орбитите на новооткритите комети и астероиди, търсене на нови такива както и изследване на физическите им характеристики. Освен под егидата на MPC се работи по съвместни проекти с множество обсерватории и организации по света (например Италия, Испания, UK и др.)
На 25.10.2005 г. новата обсерватория А79 отваря очите на света за ново космическо тяло- астероида 2005 UT12. Международният астрономически съюз признава новият астероид в Слънчевата система който групата български астрономи засичат. Откритието вече е вписано в каталога на световната организация. Пътят към откритието на Елина Михайлова, Чавдар Калдиев и техният ръководител Филип Фратев минава през експедиция на Юндола и 7 наблюдателни нощи там.
2005UT12
Още в първите дни след намирането на небесното тяло, директора на MPC професор Брайън Марзден потвърждава неговото съществуване. Астероидът е открит в съзвездието Бик и обикаля между орбитите на Марс и Юпитер с период от 4, 2 земни години.
Орбита на 2005UT12
Как стана самото окткритие:
Ф. Фратев:
”Един приател ми казваше, че постоянството и усилията винаги са възнаградени. Мился, че е прав. Гледахме 33 комети за около 2 месеца и половина, преоткрихме 3, установихме и преномерирахме последните ни извести периодични комети в Слънчевата система (171/P и 173/P) и т.н. В крайна сметка Октомври се оказа щастлив! По време на рутинните наблюдения на сравнително ярката 65/P Gunn (17.3м), един приятел ми звъна по телефона. От дума на дума се отнесох и снимах въпросната комета „безмислено“ много. Имено това множество от кадри в областа на еклиптиката (в съзвездието бик) ми позволи да заснема доста по-слаби обекти от предвиденото и да „хвана“ астероида!”
Няколко дена по-късно, А79 наблюдава и втори обект! Той също се оказа нов астероид, но в послесвие се оказва, че е бил наблюдаван 5 дена по-рано от англиски и американски астрономи, които в крайна сметка стават неговите откриватели. Астероида е бил засечен в близост до друга комета- P/2005 U1 и „уловен“ по идентичен начин с 2005 UT12. Поради липса на адекватно финансиране, до този момент А79 няма възможност за целенасочено търсене на нови астероиди и разчита повече на късмета, докато оползотворява наблюдателното време за определяне на орбиталните характеристики на скоро открити обекти”
До 07/01/2006 астероида беше наблюдаван 74 пъти от множество обсерватории по света. Междувременно, специалистите от А79 идентифицираха небесното тяло на архивни изображения от големия американски проект ”NEAT”. По този начин орбиталните параметри на астероида бяха определени много точно и несигурността в положението на астероида за 50г. напред е не по-вече от 1.5-2.0 arc sec, което е нищожно малко и той не може да бъде загубен от полезрението на астрономите. MPC му даде параметър на несигурност U=3 (скалата е от 1 до 10 като най-точно са определени орбитите с U=1). За този период астероида не беше идентифициран с никой познат от миналото такъв и няговото постояно наименование остава 2005 UT 12. След провеждане на нблюдения при още две сближения с Земята (2 опозиции), то той ще бъде окончателно номериран и ще му бъде дадено име. Права върху неговото кръщаване имат само Ф.Фратев и неговите колеги.
За 2005 г. IAU класира А79 на 21 място по общ брой наблюдения, а сумарния брой на нашите наблюдения и тези проведени от НАО ”Рожен”-БАН (MPC 071) поставиха България в челната десетка в тази област на науката за 2005 г.
2006 г. Продължението…
Работата по предварително подготвена програма А79 продължи и през настоящата 2006 година, но поради изчерпаме на финансовите средства това стана от пролетта на 2006г. Първоначално вниманието беше насочено към близко приближилата и разпадаща се комета 73P, като по официални данни А79 е втората обсерватория която регистрира наблюдавания по-късно дори от космическият телескоп „Хъбъл” разпад на 73P-b. От 22 юли A79 започна програма за целенасочено търсене на нови астероиди. Целта беше да бъдат засичани сравнително ярки обекти за да може тяхната орбита да бъде определяна точно, което се вписва с останалите цели на обсерваторията. „ По света повечето любители избягват да се конкурират за обектите, които са е полезрението на големите американски проекти като LINEAR, NEAT, Catalina и др. Те предпочитат да наблюдават по-слаби обекти с което си „гарантират” правата за тяхното откритие, но получения брой на наблюдения и съответно орбитата им е много неточна и води много често до тяхното безвъзвратно загубване. Това не носи научна стойност и не може да е основна задача на екип, който в голямата си част е съставен от професионалисти.”- коментира ръководителят на А79.
За 12 нощи бяха сканирани предварително определени от площадки около еклиптиката. Полето на кадър беше около 30 по 20 арк мин. За стак ( сумиране на изображения) от 15 кадъра се достигаше без проблем около 20.5 звездна величина на височина от 20-30 градуса над хоризонта. Ето и част от резултатите:
-Открити бяха три нови астероида.
2006 ОК14 е новият астероид открит от българи, а в случая на останалите два (2006 ОL14 и 2006 OE14) хората от А79 се водят като съоткриватели поради допуснати процедурни грешки при изпращане на данните. Най-общо казано според сегашната орбита, новия астероид е типичен представител на централната част на главния астероиден пояс. Той обикаля между орбитите на Марс и Юпитер с период от 4.09г. и има размер от около 3 км. По-време на откритото, астероида беше изключително ярък за астрономическите представи в областта -17.4м. Такива открития са голяма рядкост в днешно време. „Най – общо казано почти невероятно е астероид от 17.4 звездна величина да не е засечен до сега от големите проекти като ”NEAT” като се има в предвид и автоматизираното детектване на обектите при тях” –коментират от А79.
2006ОК14
Астрономите в НАО ”Рожен” начело с Веселка Радева-БАН и любители от форум ”Звездно Общество” също се включиха активно в наблюденията на астероида, което спомогна за определяне на неговата първоначална орбита. Това е много важно при ранните етапи от този вид наблюдения- астрометрия. Астрометрията е определянето на видимото движение на малките тела в Сл. Система спрямо заобикалящите ги звезди, които са неподвижни за голям период от време. На базата на множество наблюдения (на теория по три) от различни дни може да се определи орбитата на тялото, т.е неговото пространствено положение в Сл. Система. Трябва да се отбележи доброто сходство на изчислената орбита за 2006 OK14 от екипа на А79 и тази на МРС. В бъдеще IAU ще изисква освен наблюдения, всяка обсерватория да предоставя и изчислена орбита за да защити откритието си. До сега с това изцяло се занимаваше MPC. За период около един месец А79 проведе множество наблюдения на новоткритият астероид. Нашите астрономи намериха небесното тяло на изображения от 2002г. и благодарение на помоща на MPC, до момента астероида е гледан общо 105 пъти в 3 опозиции и орбитата му беше определена изключително точно. Наскоро астероида беше засечен и на стари фотографски плаки от 1953г, което ще осигури още една опозиция, което се изисква от IAU преди да му дадено име. От А79 се надяват, че това ще стане до месеци.
Едиността при наблюденията на 2006 ОК14 от двете български обсерватории доведе до задълбочаване на сътрудничеството и формиране на общи приоритети в бъдеще. България е страна с традиции в тази област на астрономията, която е изключително популярна в чужбина и ни носи голям престиж.
Освен по-горните резултати от насоченото търсене на нови астероиди бяха още:
-Наблюдавани над 70 астероида и 23 комети.
-Преоткрити бяха 2 комети, като техните ранни наблюдения доведоха до съществени уточнения на физическите им характеристики.
-Изпратени до MPC бяха над 480 астрометрични наблюдения на вече известни астероиди и комети от които 30% са близко земни (NEO). За сравнение: през цялата 2005, НАО „Рожен” и А79 пращат по около 550. На 02 и 03/09/2006г. бяха открити още два нови астероида от A79…
За разлика от предните два астероида, които бяха типични представители на вътрешната част на главния астероиден пояс (зона 2), то новите обекти имат интересни орбити. 2006 RJ беше открит през нощта на 01 срещу 02/09/2006 като обект от 18.2 м. Периода му е от 5.48г. За период само от 10 дена бяха получени достатъчно на брой наблюдения за да бъде открит на стари изображения от екипа на А79 и свързан с други такива от MPC. В следствие на това на астероида беше получена точно определена орбита като небесното тяло е от крайната-външна част на главният пояс (зона 3). Наблюденията на този астероид с абсолютна звездна величина от над 14,6м и приблизителен диаметър от 7.2км са много важни, защото само по този начин ще може да се определи в кой под орбитален клас ще попадне, а от там пък произтичат и ред други неща. За момента той попада в класа на Hygiea астероидите, които до май 2004 бяха само 1703 на брой от общо над 214 000 известни в Слънчевата система. Нито един от този клас астероиди все още не е получил окончателен номер и име, което е също предизвикателство за А79 в момента. В непосредствена близост е класа Themis, за който на скоро в ”Science” излезе публикация с хипотеза за трети пояс/облак от комети. Астрономите на хавайските обсерватории провеждат интензивно търсене на такива слабо активни комети в главния астероиден пояс, които вероятно отдавна са покрити с дебела прахова кора.
2006RJ
2006 RQ е астероид с диаметрално противоположни орбитални параметри в сравнение с 2006 RJ. Периода му е около 3.37г. и той е представител на най-вътрешната зона-1 от пояса. В този случай също трябва да се уточнява орбитата на астероида с размер малко над 1км. Проблемът тук е, че астероида е много слаб (19.3R) и няма да превиши 19м, което прави наблюденията му изключително трудни. Малко може да се разчита и на големите проекти и варианта е само ние да го наблюдаваме – казват от А79 , а при липсата на средства което е основното ще търсят помощ за наблюденията си от приятелите в чужбина.
Екипа на А79 за момента включва Филип Фратев, Елина Михайлова, Чавдар Калдиев и Радослав Станчев.
Тези толкова познати ни птици, всъщност, предизвикват повече въпроси, отколкото отговори. Що е то кръгов вид? Що е то вид изобщо? Как еволюира той? Въпроси с много трудни отговори.
Кръговите видове – прав ли е Дарвин?
През 1859 г излиза от печат „Произходът на видовете” на Чарлз Дарвин. Той публикува своя труд след като 18 години го е държал в „чекмеджето”, образно казано не е посмял да даде публичност на едно толкова авангардно мислене в областта на биологията във време, когато се е считало че единствен Господ Бог е сътворител на света и всичко живо в него.
Днес, макар да са изминали 154 години от отпечатването на този епохален труд, променил човечеството, макар вече да не царува догмата за божествения произход, все още не спират споровете относно описаният от Дарвин път за поява на видовете.
Описването на същесвуващи в природата,така наречени кръгови видове отново отваря въпроса за произхода на видовете.
Какво е кръгов вид?
Застъпваща се серия от популации на близкородствени видове, при които на всяка от две съседни, видовете се чифтосват помежду си и дават плодовито потомство, но съществуват поне две и то крайни популации, при които видовете са толкова различни, че не могат вече да се чифтосват помежду си. По този начин съществува генетичен поток между съседните популации. В двата края популациите или са твърде близко географски или се застъпват, така че цялата серия описва кръг.
Различно оцветените фигури представят различните популации, като всеки две съседни се припокриват географски и слабо се отличават по условия , начин на живот, външни белези / фенотипно и генотипно/. Това разположение на популациите може да е градуално – например в планините /фиг А/ или да описва кръг около даден воден басейн /фиг В и С/. Видовете от съседните популации могат да се чифтосват помежду си, но тези от популациите в двата края / на фиг зелено и цикламено / вече са толкова различни, че не могат. Трябва да се отбележи също, че тези популации не отговарят на логиката на преносимост на свойствата и ако А популация се чифтосва с Б популация , а Б със С, то това не означава, че А се чифтосва с С, а напротив.
Кръговият вид – доказателство за еволюцията?
Ричард Докинс е най-популярния защитник на нео дарвинизма в съвремието ни и в своя труд The Ancestor’s Tale (Разказът на прародителя) (2004), отбелязва за кръговите видове, че те „ единствено ни показват в географско измерение, какво се е случило във времево.”
Докинс и редица други автори разглеждат кръговите видове като доказателство в съвремието ни за еволюцията описана от Дарвин. Като във времеви план, живите и описани днес видове са крайни популации на кръгови видове, съществуващи някога, на които междинните популации са изчезнали във времето. С други думи кръговите видове ни дават доказателство за еволюцията, като илюстрират какво се е случило през времето когато популациите генетично дивергират и постепенно се достига до появата на нов вид.
Разбира се има и немалко противници на тази теза и на практика дори научни разработки, които оборват първо примерите които ще изредим дали са кръгови видове и с това поставят основата на отричане на тази теория като цяло.
Кръговите видове поставят казуса за т.нар видов проблем, за тези които искат да разделят живите организми на точно определени и строго дефинирани видове и таксони. Оказва се, че точно при кръговите видове съседните популации е трудно да бъдат определени като отделни видове и в повечето случаи са определяни като подвидове на един вид и по- късно отделяни в отделен токсон, което пък от своя страна е оспорвано. В тези случаи единствено двете крайни популации твърдо са определяни като отделни видове. По този начин е трудно да се определи дали кръговите видове реално са представлявани от популации на подвидове или са представлявани от популации на близки, но различаващи се видове. С това се поставя отново проблема за определянето на вида като цяло. Кръговите видове в природата
Общо четири групи видове са разглеждани като примери за кръгови видове. Класически пример са чайките от род. Larus, които през 1925г. се описват ,че образуват верига от вариетети около Полярния кръг / макар по последни изследвания въпроса за това дали чайките са кръгов вид е спорен/. През 1938 се залага хипотезата за кръгово разположение на популациите на Зеленикавия певец / от сем.Коприварчета/ разпростарен в Непал около Тибетското плато, достига до Сибир, където крайните популации вече не се чифтосват помежду си. Пойното врабче Melospizamelodia заобикаля пустинята Сиера Невада в Калифорния. Саламандрите от род Ensatina формират кръг около района на Центръл Вали в Калифорния.
Тези описани случаи, както и някой други открития способстват Ернст Маер първи да формулира теорията за кръговите видове през 1942 . Съвсем наскоро през 2012 е описан и първия кръгов растителен вид.Euphorbiatithymaloides, вид млечка, образуваща кръг от Централна Америка, Карибите, който се затваря при Вирджинските острови.
Чайките – класически кръгов вид
Сребристата чайка (Larus argentatus) (отпред) и Малката черногърба чайка (Larus fuscus) (отзад) в Норвегия : два фенотипа с ясни различия
Класически пример за кръгов вид са чайките от род Larus, чиито популации оформят кръг около Северния полюс. Популациите са определени като отделни видове. Европейската сребиста чайка (L.argentatusargenteus), която е разпространена на територията на Великобритания и Ирландия може да хибридизира с Американската сребриста чайка (L. smithsonianus), която обитава района на Северна Америка. Тя от своя страна може да хибридизира с чайката вега или източно сибирската сребриста чайка (L. vegae). При западните подвидове чайката на Бирула (L. vegaebirulai), хибридизира с чайката на Хюглин (L. heuglini), която от своя страна хибридизира със сибирската малка черногърба чайка (L. fuscus). Четири от гореспоменатите видове обитават северната част на Сибир. Последният е източен представител на малка черногърба чайка ( (L. fuscus) распространил си в северо-западна Европа, включително Великобритания.
Малката черногърба чайка и сребристата чайка са достатъчно различни, за да не хибридизират нормално / не се чифтосват дори при изключителни обстоятелства/ .
Така чайките оформят непрекъсната серия от популации с изключение, където двете линии се срещат в Европа.
Въпроси с неокончателни отговори
Кръговите видове отварят редица фундаментални въпроси, на които все още не е даден окончателен отговор.
Как се определя вида?
Какъв е начинът на поява на нов вид?
Кръговите видове доказателство за еволюция ли са?
Защо са описани толкова малко случаи на кръгови видове?
Какво може да ни помогне наблюдението и изследването на кръгови видове по отношение на опазването на видовото разнообразие?
На тези и редица други въпроси отговорите предстоят.
^ Liebers, D.; De Knijff, P.; Helbig, A. J. (2004). „The herring gull complex is not a ring species“. Proceedings of the Royal Society B: Biological Sciences271 (1542): 893.doi:10.1098/rspb.2004.2679. edit
Moritz, C., Schneider, C.J., et al. (1992). „Evolutionary relationships within the Ensatina eschscholtzii complex confirm the ring species interpretation“. Systematic Biology41: 273–291.
Дълги години семейството има оспорвана систематика и видовете са отнасяни като подвидове и обратно. Учените от Дружеството на британските орнитолози/ BOURC Taxonomic Sub-committee /, уточняват окончателно приетата таксономия на сем. Laridae
/J. Martin Collinson, David T. Parkin, Alan G. Knox,George Sangster and Lars Svensson. Species boundaries in the Herring and Lesser Black-backed Gull complex. British Birds 101 • July 2008 • 340–363/
Филогенетично дърво на чайките от сем. Laridae, основано на ДНК анализ. Това дърво се базира на изчисления за 20 000 поколения. По-дебелите черти показват вероятност по-голяма от 95%. Със син триъгълник е показано монофилията на род Larus
Ще обърна внимание само на спорните видове, с окончателно взето решение по отношение на систематиката.
• Каспийска чайка Larus cachinnans (монотипична)
Дълго време класификацията на този вид е била спорна, като е описван като общ вид заедно със жълтокраката чайка Larus michahellis и с арменската чайка Larus armenicus. Дори все още в редица сборници видът е останал с наименование жълтокрака чайка. По нови изследвания, основани на ДНК анализ, учените го определят като отделен монотипичен вид и като един от най – старите във филогенетично отношение видове, дал основа за разпространение и поява на северо западните видове чайки. • Малка черногърба чайка Larus fuscus (политипичен, със subspp. fuscus, intermedius,graellsii,heuglini, taimyrensis, barabensis)
Таксономията на групата Larus argentatus/Larus fuscus / сребриста – малка черногърба чайка/ е доста оспорвана и редица учени определят няколко подвида или вида в тази група като цяло. Освен това групата на тези птици са описани като т.нар. кръгови видове, разпространени кръгово около Арктика. Видът малка черногърба чайка Larus fuscus е определен като политипичен с няколко подвида:
Larus fuscus sbsp. Fuscus
Larus fuscus sbsp. Intermedius
Larus fuscus sbsp. Graellsii
Larus fuscus sbsp. Heuglini
Larus fuscus sbsp.taimyrensis
Larus fuscus sbsp. barabensis
• Сребриста чайка Larus argentatus ( политипичен със subspp. argentatus and argenteus)
Видът сребриста чайка Larus argentatus e определен като политипичен с два подвида:
Larus argentatus sbsp. argentatus / Скандинавска сребриста чайка/
Larus argentatus sbsp. Argenteus /Британска сребриста чайка/
• Жълтокрака чайка Larusmichahellis(политипичен със subspecies michahellis, atlantis и потенциално други популации, които тепърва се изследват/
Дълго време класификацията на този вид е била спорна, като е описван като подвид на сребристата чайка Larus argentatus или на каспийската чайка Larus cachinnans. Името си получава от немския учен Карл Михахелес /Karl Michahelles /. Някой я именуват западна жълтокрака чайка, за да я отделят от източната популация Larus atlantis , отделена като подвид Larus michahellis sbsp. Atlantis
Larusmichahellis sbsp. michahellis
• Американска сребриста чайка Larus smithsonianus (политипичен със subspp. smithsonianus,vegae,mongolicus)
Този вид също дълго време е бил спорен. Сега се према, че е отделен вид, но политипичен, като включва подвидове
Larus smithsonianus subspp. smithsonianu
Larus smithsonianus subspp. vegae
Larus smithsonianus subspp.Mongolicus Последният подвидMongolicus се разглежда от повечето автори като отделен вид. На този етап се према като подвид, но неговото положение е възможно да се промени в резултат на следващи изследвания.
• Арменска чайка Larus armenicus (монотипичен)
Дълго време класификацията на този вид е била спорна, като е описван като общ вид заедно със Каспийска чайка Larus cachinnans. Сега окончателно се према видът за отделен монотипичен – Larusarmenicus
Larus michahellis, Larus smithsonianus, Larus argentatus, Larus cachinnans, Larus fuscus техните подвидове са наричат от орнитолозите комплекс херингови чайки.
Фенотипични вариации в комплекса херингови чайки. (a) L. michahellis,
Лесбос, Гърция (b) L. smithsonianus, Нюфаунленд, Канада (c) L. a. argentatus, Ротердам, Холандия (d) L. a. argenteus, Лейден, Холандия (e) L. hyperboreus, Свалбард, (f)L. hyperboreus, Чърчил, Канада (g) L. marinus, Грейфсвалд, Германия (h) L. fuscus, Лейден, Холандия Leiden,
Литература:
The Herring Gull Complex (Larus argentatus -fuscus – cachinnans) as a Model Group for Recent Holarctic Vertebrate Radiations.Dorit Liebers-Helbig, Viviane Sternkopf, Andreas J. Helbig{,and Peter de Knijff. Springer-Verlag Berlin Heidelberg 2010
J. Martin Collinson, David T. Parkin, Alan G. Knox,George Sangster and Lars Svensson. Species boundaries in the Herring and Lesser Black-backed Gull complex. British Birds 101 • July 2008 • 340–363
Херинговите чайки – филогеография и еволюция.
Приблизитело 20 вида чайки, разпространени в Евразия и Северна Америка са наричани Херингови чайки. Таксономията на херинговите чайки е доста оспорвана. Повечето видове са описани като кръгов вид / поредица от застъпващи се географски , чифтосващи се помежду си популации, като първата и последната са толкова различни, че не се чифтосват и не дават хибриди; кръговите видове се дават за пример на пряко наблюдение на еволюцията и появата на нови видове в действие/.
За да се изясни таксономията и да се отговори на въпроса кръгов вид ли са наистина чайките, в последните години учените усилено работят върху ДНК изследване / митохондриална ДНК и полиморфни автозомални локуси ;вж. използвана литература/ на тези видове. В следствие на тези изследвания Дружеството на британските орнитолози/ BOURC Taxonomic Sub-committee / приемат единна систематика на херинговите чайки / Larus argentatus, Larus fuscus, Larus michahelis, Larus cachinnans, Larus armenicus, Larus smithsonianus, Larus mongolicus, Larus barabensis и подвидове/.
На фиг 1: Ареал на разпространение на херинговите чайки.
На основа на ДНК изследванията се изяснява и филогеографията на тези видове. Оборва се първоначалната версия, че всички тези чайки са произлезли от един общ предшественик, обитаващ определен ареал / рефугиум / , от който постепенно по кръгов начин в течение на времето се е разпространил в цяла Холарктика, давайки основа за появата на всички тези нови видове.
Въз основа на ДНК изследванията, учените отделят два ясно генетично различни клона, които преди около 300 000 години са съществували в два региона / рефугиума/ – Северно Атлантически и Арало Каспийски клонове. Каспийската / Larus cachinnans /и сребристата чайка / Larus argentatus/ са два съвременни вида, които имат най-различаващия се и най-ранно отделен от хипотетичния прародител хаплотип. На база на типизирането, учените определят, че хипотетичния прародител на първия клон е обитавал северо източен Атлантик / сегашния ареал на сребристата чайка /, а на втория клон в Арало Каспийския регион / сегашен ареал на каспийската чайка/.
Фиг 2. Филогеография на херинговите чайки – генетично обособени / на основа на мтДНК хаплотипове/ в два клона – Северно Атлантически и Арало Каспийски.
Арало Каспийската популация /предшественик на клон II/ се е разпространил до северно Евразийските брегове, след това на запад до Великобритания и Исландия / сегашния ареал на малката черногърба чайка Larus fuscus/ и на изток по цялото крайбрежие на северен Сибир / vegae, schistisagus/ и Северна Америка /smithsonianus, glaucescens, glaucoides/. Фиг 3 Филогеография на херинговите чайки – генетично обособени / на основа на мтДНК хаплотипове/ в два клона – Северно Атлантически и Арало Каспийски.
Това разпространение е в съгласие с теорията на кръговите видове, като прилежащите хаплотипове потвърждават наличието на генетичен поток между видовете, отново в съгласие с тази теория. Но липсата на колонизация на Larus smithsonianus от Северна Америка, обратно в Европа, с което да се затвори кръга е в противоречие с тази теория, тъй като краищата на описания кръг не се застъпват.
В противоречие на теорията за кръговия вид е наличието на втори клон – Северно Атлантическия – клон I, предшественика на койта дава появата на атлантическата чайка Larus atlantis , средиземноморската жълтокрака чайка Larus michahelis и арменската чайка Larus armenicus, които носят съответния хаплотип.
При Северно Атлантическия клон I , разпространението и обособяването на новите видове е вървяло на две вълни. При първата вълна на колонизация, предшественик, най-близък до атлантическата чайка Larus atlantis, навлиза към Средиземноморието и Черно море и дава основа на обособяване на арменската чайка Larus armenicus, който при последващо заледяване остава изолиран. При втората вълна атлантическата чайка дава появата на жълтокраката чайка Larus michahelis , която вторично среща арменската в източното Средиземноморие . Генетично атлантическата и арменската чайка имат по близък генотип до предшественика, отколкото жълтокраката чайка.
Фиг. 4 Пиктографско представяне на генетичните връзки между видовете херингови чайки. Линиите представят генетичната дистанция, като дължината на линиите грубо представят генетичните мутации, даващи разлики между видовете. Оранжевият кръг, предстявщ Сибирско/Американските видове включваL. glaucoides, L. glaucescens, L. hyperboreus, L. schistisagus, L. smithsonianus, L. taimyrensis, L. mongolicus,L. vegae и L. heuglini.
При тези генетични изследвания е установено най-голямо различие при Larus occidentalis и затова този вид е взет като външна група. Има два вида, а именно сребристата чайкаLarus argentatus и полярната чайка Larus hyperboreus, които имат и от двата хаплотипа ДНК. Това сложно положение е в резултата на две хипотетични ситуации – първата, че тези две чайки са носители на гените на общ за двата клона предшественик, а втората по-вероятна хипотеза е ,че наличието на двата хаплотипа се дължи на вторична по-късна хибридизация на вида, която се е осъществила след отделянето на двата вида като такива и обратна колонизация и хибридизация с видове от другия клон. Като за сребристата чайка на основа на изследвания се дава по-вероятно тя да принадлежи към клон Iи вторична хибридизация с представители на клон II , да са причина за наличие на двата хаплотипа.
Генетичните изследвания през последните години дават изключителната възможност учените да надникнат в начина на еволюция и разпространение на отдлените видове, да изчислят близки и далечни връзки между видовете и да помогнат за доказването на достатъчни разлики при обособяването на отделен вид от страна на таксономите.
Не всичко е изяснено в случая с херинговите чайки. В бъдещ план има основа да се доизясняват родствените връзки и генетичния поток при отделните видове, сложното положение на сребристата и полярната чайка. Необходимо е да се изясни все още спорното положение на някой подвидове. Дали чайките са кръгов вид също остава отворен въпрос. Ясно е, че предварително описаният от Маер модел е неточен, но това не значи че може с лесна ръка да се отхвърли като теория. Следващи изследвания ще продължат изясняването на видообразуването и разпространението на видовете.
Използвана литература:
The Herring Gull Complex (Larus argentatus -fuscus – cachinnans) as a Model Group for Recent Holarctic Vertebrate Radiations.Dorit Liebers-Helbig, Viviane Sternkopf, Andreas J. Helbig{,and Peter de Knijff. Springer-Verlag Berlin Heidelberg 2010
J. Martin Collinson, David T. Parkin, Alan G. Knox,George Sangster and Lars Svensson. Species boundaries in the Herring and Lesser Black-backed Gull complex. British Birds 101 • July 2008 • 340–363
Sternkopf et al. Introgressive hybridization and the evolutionary history of the herring gull complex revealed by mitochondrial and nuclear DNA BMC Evolutionary Biology 2010, 10:348
Dorit Liebers1†, Peter de Knijff2 and Andreas J. Helbig1*. The herring gull complex is not a ring species. Proc. R. Soc. Lond. B (2004) 271, 893–901
СРЕБРИСТА ЧАЙКА Larus аrgentatus
Този вид е с изключително широк ареал на разпространение. Видът се счита като слабо засегнат в червената листа на IUCN.
Таксономичен статус
Таксономията на групата Larus argentatus/Larus fuscus / сребриста – малка черногърба чайка/ е доста оспорвана и редица учени определят няколко подвида или вида в тази група като цяло. Освен това групата на тези птици са описани като т.нар. кръгови видове, разпространени кръгово около Арктика. По последно приетата официално таксономия на тези видове от Дружеството на британските орнитолози/ BOURC Taxonomic Sub-committee /и приета от всички орнитолози по света, двата вида са отделени като отделни видове.
Видът сребриста чайка Larus argentatus e определен като политипичен с два подвида:
Larus argentatus sbsp. argentatus / Скандинавска сребриста чайка/
Larus argentatus sbsp. Argenteus /Британска сребриста чайка/
Разпространение
Сребристата чайка гнезди в по голямата част от Канада и Аляска, също така в североизточните Американски Щати, по северното крайбрежие на Европа включително на територията на Исландия, Ирландия, Скандинавия и Англия. Гнезди в северна Русия от централната и част до краен изток. В Европа е описана да гнезди и в Западна, Централа и Източна Европа. Някои популации зимуват в далечен юг и са локализирани в по голямата част на САЩ, Мексико и Централна Америка, по северните брегове на Испания и Франция, голяма част от Япония и югоизточните брегове на Китай.
Сребристата чайка е най-добре познатата чайка по бреговете на Западна Европа.
За България е по-рядък вид. Дълго време е описван като най-разпространения вид за нашата страна, поради таксономичен спор с жълтокраката чайка Larus michahellis.
Изчислено е, че популацията на сребристата чайка наброява 2,7 – 5,7 млн индивида като цяло.
Описание и екология
Северно гнездящите популации на този вид са мигриращи, а южните, въпреки че са описани като номадски , не са мигриращи.Вида гнезди от средата на Април до късен Юни в колонии от по няколко хиляди двойки.
Обитава крайбрежни и близки до брега райони, но може да навлиза покрай реките навътре в сушата, като се придържа към езера, язовири, заблатявания в близост до широки тревисти местности. Няма специфичен гнездеви хабитат, но предпочита скалисти места близо до брегове или водни басейни, недалеч от тревисти местности, острови, дюни, пясъчни брегове, тундра, ниски заблатени местности в близост до езера. Всеяден вид, който може да се храни със всичко, което го има в излишък в района на обитание. Храни се с риба, червеи, раци и други морски безгръбначни / миди, морски звезди или морски червеи/, млади и възрастни птици, яйца, гризачи, насекоми, плодове, корени, отпадъци и мърша. Изледвания са показали, че сребристите чайки могат да виждат ултравиолетова светлина. Пият солена вода и с помощта на специална жлеза, разположена над очите, отделят излишната сол.
Видът е добре познат в градските райони по крайбрежните и крайречните населени места. Гнезди по покривите на сградите. Храни се с отпадъци, гризачи, малки птички, насекоми, като е известен с това, че редовно краде храна и рови в боклука.
Сребристата чайка е голяма чайка. Размерът варира от 60 до 67 cm. / измерено от върха на опашката до върха на клюна/. Размах на крилете 125 до 155 cm и тежина от 1,050 до 1,525 g. Женските са по-малки с размер от 55 до 62 cm. / измерено от върха на опашката до върха на клюна и тежина от 710 до 1,100 g. Размер на крилете при сгънато положение 38,1 до 48 cm, човка 4.4 до 6,5 cm и стъпало / тарзус/ 5.3 до 7.5 cm. Възрастното оперение е сив гръб и горната част на крилете, бяла глава и долната част на тялото. Краищата на крилете са черни с крилни огледала / бели петна/. Човката е жълта с червено петно. Покрай очите има бледо жълт кръг. Краката обичайно са розови, но при някой популации в Балтика, могат да бъдат бледо жълти.
Този вид е с изключително широк ареал на разпространение, като не е уточнен. Видът се счита като слабо засегнат в червената листа на IUCN. Таксономичен статус
Дълго време класификацията на този вид е била спорна, като е описван като общ вид заедно със жълтокраката чайка Larus michahellis и с арменската чайка Larus armenicus. Дори все още в редица сборници видът е останал с наименование жълтокрака чайка. По нови изследвания, основани на ДНК анализ, учените го определят като отделен монотипичен вид и като един от най – старите във филогенетично отношение видове, дал основа за разпространение и поява на северо западните видове чайки. Разпространение:
Larus cachinnans е разпространен в източна Европа, Средния Изток и северозападна Африка и централна Азия. Резидентен по крайбрежието на Черно и Каспийско море. Сезонно гнезди от Черно море на север на Каспийско море до източен Казахстан и азиатските степи. Зимува по бреговете на юго западна Азия, северо западните брегове на Африка и около Арабския полуостров нагоре до северозападна Индия. В Европа се среща до Полша и източна Германия, на север Швеция, Дания и Норвегия. Дори са наблюдавани и в Англия. Срещани са до Червено море и Персийския залив.
Постоянно пребиваващ: Авганистан, Армения, Босна и Херцеговина, България, Китай, Джибути, Египет, Етиопия, Грузия, Индия, Иран, Ирак, Израел, Йордания, Казахстан, Кувейт, Ливан, Оман, Пакистан, Палестина, Катар, Румъния, Суадитска Арабия, Сомалия, Сирия, Турция, Украйна, Обединените Арабски Емирства, Йемен
Временно пребиваващ: Бахрейн, Малдиви, Непал и Шри Ланка
Kлючови факти
Текуща IUCN Red List категория
Слабо засегнат
Семейство
Laridae Чайки
Първи описан
Pallas, 1811
Популация
Разширяваща се
Ендемит
Не
Описание и екология
Видът е изцяло мигриращ, макар някой колонии в Каспийско и Черно море да се считат за постоянни. След гнездене миграциите за зимуване са от юли до ноември. За гнездене се завръщат от средата на февруари до средата на юни. Гнездят от средата на март до април, макар точният период да варира географски. Гнездят в колонии от повече от 8000 двойки и могат да образуват групи и в смесени колонии.
Гнездят близо до реки и речни корита с обрасли наноси, заблатявания, тревисти влажни местности, скални морски пукнатини и пясъчни брегове, речни и морски острови, каменисти брегове, дюни, бракични крайбрежни заблатявания, обрасли с треви и храсти. Извън периода на гнездене обитават крабрежието и навлизат навътре по реките, като често се хранят и с култивирани посеви.
Всеяден вид. Менюто им включва риба, безгръбначни / насекоми, мекотели, раци/, влечуги, малки бозайници / полевки, катерици, лалугери/, боклук, мърша, яйца, пилета, осттатъци от храна.
Каспийската чайка е голяма чайка. Размерът им варира от 56 до 68 cm. / измерено от върха на опашката до върха на клюна/. Размах на крилете 137 до 155 cm и тежина от 680 до 1,590 g. Размер на крилете при сгънато положение 38,5 до 48 cm, човка 4.6 до 6,4cm и стъпало / тарзус/ 5.8 до 7.7 cm. Каспийската чайка има дълга слаба човка, завършваща в наклоненото чело. Краката, крилете и гърба са по-дълги от на сребристата и жълтокраката чайка. Очите са малки и често тъмни, краката варират от бледо розови до бледо жълтеникави. Гърба и крилете са слабо по-тъмно сиво от това на сребристата чайка, но слабо по-бледо сиви от на жълтокраката чайка.
МАЛКА ЧЕРНОГЪРБА ЧАЙКА Lesser Black – backed Gull Larus fuscus
Този вид е широко разпространен. Видът се счита като слабо засегнат в червената листа на IUCN.
Таксономичен статус
Таксономията на групата Larus argentatus/Larus fuscus / сребриста – малка черногърба чайка/ е доста оспорвана и редица учени определят няколко подвида или вида в тази група като цяло. Освен това групата на тези птици са описани като т.нар. кръгови видове, разпространени кръгово около Арктика. По последно приетата официално таксономия на тези видове от Дружеството на английските орнитолози/ BOURC Taxonomic Sub-committee /и приета от всички орнитолози по света, двата вида са отделени като отделни видове.
Видът малка черногърба чайка Larus fuscus е определен като политипичен с няколко подвида:
Larus fuscus sbsp. Fuscus
Larus fuscus sbsp. Intermedius
Larus fuscus sbsp. Graellsii
Larus fuscus sbsp. Heuglini
Larus fuscus sbsp.taimyrensis
Larus fuscus sbsp. barabensis
Разпространение:
Гнезди в централна и северна Русия, Скандинавия, Германия, Белгия, Холандия, северна Англия към Исландия. Гнезди целогодишно по бреговете на Португалия, южна Ирландия, Англия и северна Франция. Сезонно гнездещи популации са срещани и североизточна Испания. Сезонни гнездещи популации са широко срещани, разширяващи ареала по цялото крайбрежие на Северно море, Средиземноморието, Черно море и бреговете на Каспийско море, северните и източните брегове на Африка / включително и прилежащите реки/ и от Арабския полуостров до северозападна Индия.
Като цяло вида е с разширяващ се ареал на местообитание и гнездене. Включително и разширяване към и в Северна Америка.
Описание:
Голяма чайка / много прилича на сребристата Larus argentatus и голямата черногърба чайка Larus marinus / с големина на тялото от върха на клюна до края на опашката 51–64 cm, разстояние между крилете в полет 124–150 cm и тежина 452–1,100 g. Мъжките са малко по-големи от женските. Криле в прибрано положение / ъгъл 90’/ 38.3 to 45 cm, човка 4.2 to 5.8 cm и стъпало / тарзус / 5.2 to 6.9 cm. Спрямо видовете, с които често се бърка, малката черногърба чайка е малко по-малка и по-слабо телосложение. С жълти, рядко розови крака и малки бели крилни огледала в края на всяко крило. Възрастните има черен или тъмно сиви крила и гръб. Човката е жълта с червено петно, което младите кълват като сигнал за хранене.
Поведение:
Повечето популации са мигриращи като се местят скокообразно от място на място. Есенната миграция започва от негнездещи птици през късен Юни, последвани от гнездещите през късен Юли до Септември. Завръщането на птиците по гнездещите места започва от Февруари до ранен Юни. Птичите колонии са от смесен тип / често със сребристата чайка/ и наброяват от няколко двойки, до десетки, стотици и дори хиляди индивида.
Извън размножителния период видът остава общителен и мигрира на групи под 10 индивида.Обикновенно се храни на групи от стотици индивиди по сметища или плитчини с риба.
Хранене:
Видът е всеяден. Менюто на чайките включва малки рибки / най вече атлантическата херинга Clupea harengus, водни и сухоземни безгръбначни / бръмбари, мухи и техни ларви, мравки, пеперуди, скакалци, ракообразни, мекотели, прешленести червеи и морски звезди/, яйца на други птици и малки новоизлюпени птичета, мърша, карантии, гризачи, плодове и зърнени храни
ЖЪЛТОКРАКА ЧАЙКА Yellow – legged Gull Larusmichahellis
Този вид е с изключително широк ареал на разпространение. Видът се счита като слабо засегнат в червената листа на IUCN.
Таксономичен статус
Дълго време класификацията на този вид е била спорна, като е описван като подвид на сребристата чайка Larus argentatus или на каспийската чайка Larus cachinnans. Името си получава от немския учен Карл Михахелес /Karl Michahelles /.Някой я именуват западна жълтокрака чайка, за да я отделят от източната популация Larus atlantis , отделена като подвид Larus michahellissbsp. atlantis. Разпространение: Larus michahellis е разпространен в Европа, Средния Изток и западна Африка. Резидентен за цяла Южна Европа, по бреговете на Средиземно море, Черно море, Каспийско море, Азорите и Мадейра, Португалия, Канарските острови, Испания. Зимуват по бреговете на югозападна Азия, Европа нагоре от Германия и от Африка до източното Средиземноморие.
Описание и екология
Популациите могат да бъдат установени или разпръснати. Миграциите за зимуване са от юли до ноември и се завръщат за гнездене от средата на февруари до средата на юни. Гнездят от средата на март до април. Гнездят групово в колонии от по 8000 двойки и могат да създават групи в смесени иначе с други видове колонии.
Гнездят близо до реки и речни корита с обрасли наноси, заблатявания, тревисти влажни местности, скални морски пукнатини и пясъчни брегове, речни и морски острови, каменисти брегове, дюни, бракични крайбрежни заблатявания, обрасли с треви и храсти. Извън периода на гнездене обитават крабрежието и навлизат навътре по реките, като често се хранят и с култивирани посеви.
Всеяден вид.Менюто им включва риба, безгръбначни / насекоми, мекотели, раци/, влечуги, малки бозайници / полевки, катерици, лалугери/, боклук, мърша, яйца, пилета, осттатъци от храна.
Жълтокраката чайка е голяма чайка. Размерът им варира от 52 до 68 cm. / измерено от върха на опашката до върха на клюна/. Размах на крилете 120 до 155 cm и тежина от 550 до 1,600 g. Размер на крилете при сгънато положение 40.8 до 47.2 cm, човка 4.6 до 6 cm и стъпало / тарзус/ 5.6 до 7.5 cm. Възрастните са изключително близки по външен вид до сребристата чайка Larus argentatus, но са със жълти крака, което е причина в миналото да бъдат описани като подвид на сребристата чайка. Имат сив гръб, леко по-наситено сив от този на сребристата чайка, но по-светъл от този на Малка черногърба чайка Larus fuscus. Те приличат на последната по белочелието през есента, и черните пера с бели огледала. Имат червено петно на човката, както целия комплекс чайки. Имат и червен пръстен около очите, подобно на Малка черногърба чайка Larus fuscus и за разлика от сребристата чайка, при която този пръстен е наситено жълт.
Видове чайки от род LARUS, описани на територията на Балканския полуостров
МАЛКА ЧЕРНОГЛАВА ЧАЙКА Mediterranean gull Larus melanocephalus
Дължина на тялото:37 см; Размах на крилете:96 см.
Лете възрастните са бели с черна качулка на главата, сив гръб и сиви крила. Клюнът и краката са яркочервени. През останалото време цялото оперение е бяло, но с тъмно петно зад окото. Младите отгоре са сивокафяви с черни първостепенни махови пера и черна ивица на края на опашката. Зиме възрастните се отличават от тези на речната чайка по окраската на главата и бялата долна част на крилата, а младите- по оцветяването на първостепенните махови пера. Рее се повече от другите видове дребни чайки. Издава пронизителен крясък „ кееер”, подобно на рибарка. На Балканския полуостров обитава пясъчни коси в различно солени водоеми; по време на миграции и през зимата се среща по морски крайбрежия и навътре в сушата. Най-много популации са описани на територията на Гърция и Албания. Описан е като уязвим вид в Червената книга на България.
РЕЧНА ЧАЙКА Black-headed Gull Larus ridibundus
Дължина на тялото: 36см; Размах на крилете: 105 см.
Възрастните през размножителния период наподобяват тези на малката черноглава чайка, но качулката на главата е забележимо по-малка и първостепенните махови пера отдолу са черни. Клюнът и краката са яркочервени. Възрастните зиме и младите наподобяват малката черноглава чайка. Издават силни и пронизителни крясъци. Гнезди в Евразия от Исландия на изток до п-в Камчатка. На Балканския полуостров обитава сладководни блата и езера с обилна растителност; гнезди на шумни колонии. По време на миграции и зиме се среща в разнообразни водоеми, сметища, рибарници и др. Описана като застрашен вид в Червената книга на България.
Дължина на тялото: 43см; Размах на крилете: 105 см.
Лете възрастните са със сиви крила с черни върхове; останалото оперение е бяло с розов отенък по гърдите. Зиме розовият отенък липсва, а зад окото се появява тъмно петно. Младите са със сиво-бяло оперение с черни върхове на първостепенните махови пера. Наподобява речна чайка в извънбрачно оперение, от което се отличава по едрите размери, забележимо по-дългия клюн и оцветяването на крайните първостепенни махови пера / при дългоклюната чайка 5-6 са с черни върхове, а при речната-7-8/. Мълчалива чайка. Гнездовият ареал е Гърция, Мала Азия и северното крайбрежие на Черно море до Персийския залив и Централна Азия. На Балканския полуостров обитава бракични, солени или свръхсолени плитки водоеми; по време на миграции и зиме се среща по морски крайбрежия. Най-много популации са описани в Гърция.
МАЛКА ЧАЙКА Little Gull Larus minutus
Дължина на тялото: 43см; Размах на крилете: 105 см.
Зиме възрастните са сиво-бели с по-тъмни петна по главата; през всички сезони крилата отдолу са черни, по ръбовете с бяла ивица. През лятото главата е черна. Клюнът е тъмно червеникавокафяв, а краката – червеникави. Младите отгоре са сиво-кафяви с черно теме и черна ивица по крилата; наподобяват младите на трипръстата чайка, от която се различават по черните теме и плещи, не врязаната опашка и забележимо по-малките размери. Мълчалива чайка. Гнезди от Източна Европа на изток до река Лена в Русия. На Балканския полуостров по време на миграции и зиме се среща често по морските крайбрежия.
БЕЛООКА ЧАЙКА White – eyed Gull Larus leucophhtalmus
Дължина на тялото: 41см; Размах на крилете: 107 см.
Възрастните са с черна глава и гърло, сив гръб, бяла опашка; около окото има бял пръстен. Долната страна на тялото е бяла. Крилата са кафяви с черни махови пера; отгоре черни са само първостепенните махови пера. Клюнът е прав, кораловиден с черен връх. Краката са яркожълти. При младите гърбът и крилата са кафяви; средните кормилни пера са черни, а краката – светлокафяви. Издават разнообразни звуци. В Западна Палеарктика гнезди в Североизточна Африка. На Балканския полуостров се среща рядко по морски крайбрежия, регистрирана е в Гърция.
Дължина на тялото: 50см; Размах на крилете: 127см.
Възрастните са сиво – бели с черни върхове на крилата. По върховете на крайните първостепенни махови пера има малки бели петна, а около окото – червен пръстен. Клюнът е яркочервен с черно субтерминално петно. Краката са маслиненозелени. Зиме е с кафяви щрихи по главата. Младите са светлокафяви с черна ивица през второстепенните махови пера, сиво-кафява опашка с черна широка ивица на върха. Отличава се от дългоклюната чайка по късия масивен клюн и липсата на розов отенък по лятното оперение. Крясъците и наподобяват тези на гъска. Разпространена е по Средиземноморието. На Балканския полуостров гнезди на колонии по скалисти острови и скита по морски крайбрежия. Най-много колонии са описани в Гърция.
ЧАЙКА БУРЕВЕСТНИЦА Common /Mew/ Gull Larus canus
Дължина на тялото: 41см; Размах на крилете: 120см.
Възрастните зиме са със сиви крила и гръб, с черни ръбове на първостепенните махови пера и с бели петна по върховете им; темето и тилът на петна; през лятото цялата глава е бяла. Клюнът е зеленикаво – жълт с тесен черен пръстен, който през лятото изчезва; краката са зеленикави. Младите са пъстрокафяви, опашката им е бяла с широка черна ивица на върха. От сребристата и жълтоногата чайка се отличава по големина, по липса на червено петно на клюна и по зеленикавите крака. Издава пронизително „ киаа-киаа”. В Западна Палеарктика гнизди в Северозападна Европа. На Балканския полуостров мигрира и зимува често по морски и речни крайбрежия.
КАСПИЙСКАЧАЙКАCaspian Gull Larus cachinnans
Дължина на тялото: 62см; Размах на крилете: 141см.
Възрастните лете са бели със сиви плещи и крила, само външните първостепенни пера са черни с бели върхове /второто от тях има бял връх/, около окото без червен пръстен, ирисът е тъмен. Клюнът е с яркожълт цвят и червено петно на подклюнието. Краката са жълти, сивкави или бледорозови. Младите са кафяви отгоре с черни петна и бели отдолу, клюнът е черен, оперението на възрастните се появява на третата година. Гнезди по долното течение на р. Дунав и северните брегове на Черно и Каспийско море на изток от Централна Азия. На Балканския полуостров обитава речни крайбрежия и населени места.
Дължина на тялото: 55см; Размах на крилете: 130см.
Възрастните лете са бели със сиви плещи и крила, на които външните първостепенни пера са черни с бели върхове /второто от тях има черен връх/ , около окото има червен пръстен, ирисът е бледожълт. Клюнът е дебел, ярко жълт и с червено петно. Краката са жълти. През зимата главата и вратът са с кафяви щрихи. Младите са сиво – кафяви; клюнът и ирисът са черни, а краката – бледорозови. Оличава се от Larus cachinnans по червения пръстен около окото, светлия / не черен/ ирис, по-късия и по-дебел клюн и черния връх на второто първостепенно махово перо. Издава силни крясъци. Гнезди по северните и югозападните брегове на Средиземно море и Мала Азия, а на Балканския полуостров – на необитаеми скалисти острови и в градове, все по- често гнезди навътре в сушата.
МАЛКА ЧЕРНОГЪРБА ЧАЙКА Lesser Black – backed Gull Larus fuscus
Дължина на тялото: 53см; Размах на крилете: 134см.
Възрастните през зимата са бели с тъмни щрихи около окото и на врата, с черни крила и гръб.Клюнът е жълт с оранжево – червено петно при върха, а краката – бледожълти. Лете главата и вратът са без щрихи. Младите имат черен клюн, пъстрокафяво оперение отгоре и и бяло отдолу. Рядко издава силни крясъци подобно на сребристата чайка. В Западна Палеарктика гнезди в Северозападна Европа и на Пиринейския полуостров. На Балканския полуостров се среща по морски крайбрежия и открито море само по време на миграции и през зимата.
ГОЛЯМАЧЕРНОГЪРБАЧАЙКАGreat Black – backed Gull Larus marinus
Дължина на тялото: 71см; Размах на крилете: 157см.
При възрастните гърбът и горната страна на крилата са черни с бели ръбове на маховите пера; останалото оперение е бяло. Клюнът е жълт с оранжево – червено субтерминално петно на подклюнието, а краката – розово-червени. Младите през първата година са сиво-кафяви. Гърбът става черен през втората година. Отличава се от малката черногърба чайка по забележимо по-едрите размери и по цвета на краката. Издава крясъци „ аоук” или „ ук-ук-ук”. В Западна Палеарктика гнезди в Северозападна Европа. На Балканския полуостров се среща по морски крайбрежия само по време на миграции и през зимата.
МАЛКАБЕЛОКРИЛАЧАЙКАIceland Gull Larus glaucoides
Дължина на тялото: 56см; Размах на крилете: 146см.
Възрастните са бели със сив гръб и горна част на крилата; само първостепенните махови пера са бели; главата и шията са изпъстрени със светлокафяви щрихи. Клюнът е жълт с червено петно на подклюнието; около окото има слабо забележим червен пръстен. Краката са розови. Полетът е бърз и наподобява този на трипръстата чайка. Младите през първата година са светлокафяви с многобройни кафяви щрихи и препаски. През втората година оперението е по-светло с неясни пъстрини. От полярната чайка се отличава трудно по дребното тяло. Издава крясък „ киор-кор-кор”, подобно на сребристата чайка. В Западна Палеарктика гнезди в Исландия, о-в Шпицберген и Северна Русия. На Балканския полуостров скита по морски крайбрежия,, като е съобщена за Гърция.
АРМЕНСКА ЧАЙКА Armenian gull Larus armenicus
Дължина на тялото: 62см; Размах на крилете: 132см.
Възрастните и младите наподобяват много жълтоногата чайка. Възрастните се отличават от нея по черното петно, разположено пред червеното петно на подклюнието, и едно бяло петно / не две/ при върха на най-външното махово перо. Краката са яркожълти. Отличаването на младите е много трудно. Гнездовият ареал е в Централна Турция и Армения., а зимовищата – по източното крайбрежие на Средиземно море, най-вече в Израел и Северен Египет. На Балканския полуостров е регистрирана в България и Гърция.
ГОЛЯМА БЕЛОКРИЛА ЧАЙКА Glaucous Gull Larus hyperboreus
Дължина на тялото: 65см; Размах на крилете: 157см.
Възрастните наподобяват много възрастните белокрили чайки, но главата е по- едра, а клюнът – по-масивен, бледожълт с черен връх; около окото има жълт пръстен. Краката са розови. Полетът е тежък и тромав. Младите през първата година са светлокафяви, с многобройни кафяви щрихи и препаски. През втората година са по светли. Отличаването на възрастните и младите от белокрилата чайка при недостатъчно осветление е голямо разстояние е силно затруднено. През зимата и есента рядко издава крясъци. В Западна Палеарктика гнезди в Исландия и Северна Русия. На Балканския полуостров скита по морски крайбрежия, регистрирана е в Словения, България и Румъния.
ГОЛЯМА ЧЕРНОГЛАВА ЧАЙКА Pallas’s Gull Larus ichthyaetus
Дължина на тялото: 59см; Размах на крилете: 155см.
Зиме възрастните са с бяла глава и тъмнокафяви петна под очите и на тила; гърбът и крилата са сиви, върховете на първостепенните махови пера са повече черни, отколкото бели; останалото оперение е бяло. Клюнът е жълт с червено петно и черна напречна ивица на върха. Краката са яркожълти. Лете главата е черна. Младите през първата година отгоре са пъстрокафяви, а отдолу – бели с кафява огърлица. Издава подобни на жълтоногата чайка звуци. В Западна Палеарктика гнезди около Азовско и Каспийско море, както и на Кримския полуостров. На Балканския полуостров скита по морски крайбрежия.
Повече за птиците на Балканския полуостров може да намерите в полевия определител „ Птиците на Балканския полуостров” – Таню Мичев, Димитър Симеонов, Любомир Профиров; Издателство ЕКОТАН, откъдето е заимствана информацията.
Автори: Димитра Лефтерова и Чавдар Черников Източник: nauka.bg
“…Сам Дядо Коледа – дребничък стар симпатяга,
весело свиркаше, бързо подканяше впряга:
– Хайде, еленчета! Дашър и Виксен, и Прансър!
Бързайте, Дондър и Комет,и Кюпид, и Дансър!
Блитцен, светкавице моя, Напред към комина!
Знаете всички, че трябва през него да мина!
Както наесен летят с урагана листата,
тъй и елените литнаха, а след тях и шейната…”
Северен елен, карибу, северный олень , reindeer, caribou – Rangifer tarandus, Linnaeus, 1758
Субарктически и арктически елен, наречен северен в Европа и карибу в Северна Америка е световно популярен от приказката за дядо Коледа и теглената от елени негова шейна. Името rein /deer/ на викингски хрейн hreinn, която пък идва от протогермански храйназ hrainaz и прото – индоевропейски кройнос kroinos;, което означава рогато животно. При уралските езици – сами – поатцу poatsu , боатцу boazu,,боатсой boatsoj ; марийски – пуцъ pučə и удмурт – путцей pudžej ; Финското поро poro . В Рурсия северните народи го наричат сокжой. Името карибу caribou , от френски , произлязла от думата халибу qalipu, която на микмакски означава снежен копач, поради това, че карибуто разкопава снега, за да намери лишеи за храна. Както и да се наричат всички те са един вид – Rangifer tarandus. Вида има няколко подвида, съществуващи до днес, които са разнообразни по морфологични форми. Разделянето на групи на тези подвидове се основава или на географски райони: Северно американски и Евразийски подвидове; или на екологични групи: високо островни арктически, тундрови и горски.
Тундрови подвидове:
Характерно за тях е, че имат дълги, разклонени рога.
Евразийски тундров северен елен /Див северен елен/ (R. tarandus tarandus) – обитава арктическата тундра на Евразия, включително Скандинавския полуостров на Северна Европа. В Русия вида наброява около 1млн индивида.
Аляска карибу, Поркюпайн карибу, Карибу на Грант R. tarandus granti – разпространен в Аляска, Юкон и северозапдна Канада. Много сходен на R. tarandus groenlandicus. Наречен е на морско свинче, на река Поркюпайн, Porcupine River
Канадско тундрово карибу R. tarandus groenlandicus, разпространено в северозападна Канада, Нунавут и Западна Гренландия
Горски подвидове:
Характерно за тях е по- едро тяло, къси и тежки рога. 30-40% от женските са без рога, което според някои автори, това е адаптация към горския начин на живот.
Евразийски горски северен елен, Финландски горски северен елен R. tarandus fennicus – в див вид обитава два региона на Скандинавския полуостров и в Карелия. Неголяма популация обитава в центъра на южна Финландия. Карелската популация мигрира навътре в Русия, въпреки че е отворен въпроса дали източните представители спадат към този подвид.
Северноамериканското горско карибу Rangifer tarandus caribou – Първоначално подвидът е бил разпространен в северноамериканската тайга от Аляска до Нюфаунленд и Лабрадор и на юг до Нова Англия, Айдахо и Вашингтон. Горското карибу е изчезнал от по – голямата част от районите на обитаване и е обявен за застрашен вид. Изключение правят районите на Квебек и Лабрадор, Канада. В Британска Колумбия, областта Карибу е кръстена на горското карибу, макар видът да е изчезнал там през минали век. Съществува стадо защитени елени карибу в планините Карибу в Алберта.
Арктични подвидове:
Характеризират се с малко тяло и къса муцуна. Малкото тяло намалява повърхността на тялото, спрямо обема, което е присобление за живот в суровите арктични условия.
Свалбардски северен елен , Шпицбергенски северен елен R. tarandus platyrhynchus — най-малкия северен елен, обитател на архипелага Шпицберген/Свалбард в Норвегия. Понастоящем е най-малочисления подвид от всички елени.
Карибу на Пири R. tarandus pearyi, разпространен на северните острови на Нунавут и северозападните територии на Канада.
Арктически северен елен R. tarandus eogroenlandicus — преди е бил разпространен в източната част на Гренландия, но след 1900 г. се счита за напълно изчезнал.
Подвида Карибу на Даусон, Rangifer tarandus dawsoni е обитавал островите Хаида Гуай / Кралица Шарлот/. Последните документирани убити от ловци индивиди са от 1908г. На трите убити елена черепите и кожите се пазят до днешен ден в Кралския музей на Британска Колумбия във Виктория. Направени са генетични изследвания на този подвид и на съвременни северно американски подвидове, но изследванията не показват значителни генетични разлики между тях.
Последните елени карибу на Даусон, убити от ловци през 1908 на островите Хаида Гуай. Снимката е след отстреля на животните.
Северния елен e чифтокопитно млекопитаещо, което се отнася към сем. Еленови Cervidae, подсемейство Capreolinae. Живее в северните части на Евразия и Северна Америка , като се храни не само с треви и лишеи, но и с малки бозайници и птици. Северния елен е мигриращ вид и благодарение на тези миграции лишейния покров не се унищожава напълно и успява да се развие отново. 90% от менюто им включва лишеи – еленови мъхове Cladonia rangiferina, Cladonia alpestris, Cladonia sylvatica и др., които се явяват основна храна, която елените откриват и под снега. Лишейните вещества, получени от храната ги предпазват от стомашни паразити. Като всички преживни животни имат четириделен стомах. Хранейки се основно с лишеи през зимата в стомаха им се отделя ензима лихеназа, който разгражда лихенина до глюкоза. Освен лишеи понякога се хранят и с листа на върби и брези, също така различни треви. През пролетта, когато са стресирани по изключения може да се хранят и с малки гризачи като леминги, риба и яйца на птици. Някои стада на чукчите са наблюдавани да се хранят с гъби през лятото.
Широките им копита позволяват да се придвижват по рохкия сняг и да го разкопават, за да търсят храна. Жаждата си утоляват в продължение на 9 месеца в годината със сняг. Козината е топла, плътна с гъст пухест долен слой. Основните косми достигат дължина до 1-2,5 см.и са с плътност 2000 косъма на кв.см. Космите са кухи за по-добра топлоизолация, помагат и при плуване и преход през река. Долния слой е с плътност около 500 косъма на кв.см. Когато елена легне на снега, не го разтопява, защото козината му е устроена да пази топлината на тялото му. Северните елени са по своему уникални измежду своите събратя елени от сем. Cervidae. Те са изключително издръжливи при продължителен бяг, което не е изненадващо имайки предвид простора на тундрата. При този вид мъжките и женските носят рога, за разлика от другите видове елени, при които само мъжките развиват рога. Рогата на мъжките северни елени са пропорционално най -големи спрямо тялото, в сравнение с другите видове елени. Освен това рогата на северния елен са изключително елегантни. При женските, рогата са необходими за защита и конкуренция за храна и падат при раждане на малките еленчета. Елените са социални животни, които живеят на стада достигащи до 50 000-500 000 животни, които се събират по време на пролетта. Стадото се състои от отделни чифтосващи се групи от по 10 – 1000 индивида. Брачния период е най-често Октомври. Малките се раждат през май – юни след бременност от 228 дни. Най-често малките са едно до две, бозаят до 6 месеца и достигат зрялост на 2,5 – 3,5 години. Продължителност на живот до 20 – 25 години. Размерите на женските достигат 162-205 см на дължина и тежат от 80 до 120 кг. Мъжките поначало са по-големи и достигат 180-214 см на дължина като тежат 159-182 кг, макар някой особено големи екземпляри са достигали 318 кг. Подвида R. t. platyrhynchus от Свалбард е значително по – малък / феномен известен като островен джуджевизъм/ и женските тежат приблизително 53 кг през лятото и 70 кг през есента. Мъжките са приблизително 160 см дълги и около 65 кг през пролетта и 90 кг през есента. Този елен също така е и по-късокрак. Опитомените елени като цяло са по-късокраки и по-тежки, отколкото дивите им събратя. Цвета на козината им варира значително, както индивидуално, така и в зависимост от сезона и подвида. Северните популации като цяло са по-светли и по -малки, докато южните са по-тъмни и по-големи. Това добре се наблюдава в Северна Америка където северния подвид карибу на Пири е най-белия и най-малкия от всички континентални подвидове елени, докато най-южния подвид горското карибу e по-голямо и по-тъмно оцветено. Рогата на северния елен са изградени от кератин и различни протеини, но имат вкостена сърцевина. Освен, че са единствения вид елени, при които и мъжките и женските имат рога, те също са единствените на които рогата могат да регенерират. Рогата израстват направо на черепа. Те са меки и гъвкави, също са изградени от жива кръвоснабдена тъкан, затова са топли на пипане. Рогата показват доминирането и йерархията в стадото, но те имат не само тези цели. Женските запазват рогата през зимата, основно за да се защитават и конкурират за храна и ги загубват през пролетта, когато се родят малките. Мъжките ги губят в началото на зимата и им израстват отново през пролетта. Невкостенелите рога са известни в народната медицина с лечебните си свойства. Отделяния от тях екстракт, получен от рогата на северния елен се използва във фармакология в качеството си на общо тонизиращо и адаптогенно лекарствено средство. През 60 – те години под ръководството на проф. Брехман в Русия се разработва нов препарат от еленови рога „Рантарин”, първия получен от висококачествен екстракт от рога на див северен елен. Освен това се произвежда и хранителна добавка с имуностимулиращо действие „Цигапан” (продава се и в България). Хората опитомяват северните елени, като са изолирали част от стадата на дивите животни. Домашните северни елени живеят в полусвободна паша, а от дивите животни се отличават по това, че са свикнали с хората и в случай на опасност не се разбягват напосоки, а се събират накуп, надявайки се на защита от хората. От елените хората получават мляко, месо, кожа, рога, кости и също така се използват като ездитни животни. От човека, елените получават сол и защита от хищниците.
По последни генетични изследвания на мтДНК се определят най – малко два или три отделни и независими одомашнявания на северния елен от хората в източна Русия и Феноскандинавия /Норвегия, Швеция и Финландия/. Най – вероятно това се е случило преди 2-3 хил.г. Генетично почти не се откриват съществени разлики между домашния и дивия северен елен. Северния елен винаги си остава див. Естествено в неволя става по кротък, но и при най-малкото недоглеждане на стопанина и гордия син на Севера отново бяга на свобода. Дори за издояване или за да ги приберат на заслона е необходимо доста усилие от страна на стопаните. Въпреки това северните народи изцяло зависят от елените и като правило живота им минава с тях. Хората непрекъснато полагат грижа за елените. На Север се счита богат този, не у когото има много пари, а у когото има много елени.
Еленовъдството се практикува в Норвегия, Швеция, Финландия, Русия, Монголия, Китай, Аляска, Канада и Гренландия от повече от 20 различни етнически групи. Еленовъдите полагат грижи в големи територии от Арктика през последните няколко хиляди години. Тези ареали в ново време стават значими за хора индустриален интерес към петрола и газта. След падането на желязната завеса през септември 1993 г се организира международен фестивал на еленовъдите в Тромсо. Присъстват близо 360 представителя на различни еленовъдни стопанства от Норвегия, Швеция, Финландия, Русия и Аляска. Това е първия път когато хора от различни страни и нации се срещат във връзка с еленовъдството. Това събитие бележи интернационална кооперация на циркумполярните еленовъди в основаването на Световната асоциация на еленовъдите.
Карибуто в своите миграции изминава приблизително 5000 км на година, което е най-дългото пътешествие на сухоземно животно. Миграциите се извършват през пролетта и есента, като стадата се придвижват със скорост 19-55 км на ден. Максималната скорост, която достига карибуто е 60-80 км/час. Карибуто е много добър плувец и лесно прекосява големи реки и езера. При плуване достига скорост от 6,5 км/ч, а понякога и до 10 км/ч. Плътността на популацията е рядка, при подходящ хабитат до 0,5 животни на квадратен километър. Но по време на миграциите се събират на стада достигащи до 19 000 животни на кв.км. В Северна Америка елена е изключително мигриращ, като се мести от карйбрежията през лятото към вътрешността през зимата, докато в Европа видът е далеч по-седентарен. В Сибир северните елени извършват ежегодни пътешествия, пролетта от гората в тундрата, а през лятото, когато в тундрата се появяват много комари, обратно в гората. Тези преселения се извършват на големи стада. Като правило са последвани от вълци и други хищници, които нападат изоставащите, стари и болни животни. За здравите вълка не е страшен до образуване на твърда снежна кора. На нея северните елени често си режат краката и вълците се възползват от това като убиват окуцелите животни. Основни врагове в природата са кафявата и полярна мечка, вълците, скалния орел, росомаха. Много страдат от ятата комари през лятото в тундрата. Най-сериозен враг на елените се явява човека с неговата дейност.
Съвременните елени се счита че са се появили за пръв път в Евразия, Стария свят и са започнали да се разпространяват в Новия Свят, Северна Америка по време на плиоцена преди около 3,5 млн години. Най-старите намерени фосили от Rangifer в Северна Америка са отпреди 1,6 млн години/ зъби , Юкон/, по-нови са съответно от 45 500 г/ краниален фрагмент, Юкон/ и 40 600 г /рога, Квебек/. Произходът на карибу се е случил преди 80 000 – 10 000 г по време на последното заледяване и не е още добре изяснен. По време на това последно заледяване карибу е бил широко разпространен от двете страни на Лорентидите, на юг и на север. Осемте съвременни подвида се счита, че са произлезли от три изолирани рефугиума . Трите рефугиума са определени като Берингов, Евразийски и Южен. Беринговия рефугиум, простиращ се от Евразия, Аляска и Юкон, дава основа на поява на северноамериканските тундрови подвида Rangifer tarandus granti и Rangifer tarandus groenlandicus, арктичните подвидове Rangifer tarandus platyrhynchus, Rangifer tarandus pearyi, and Rangifer tarandus eogroenlandicus, евразийския тундров подвидов Rangifer tarandus tarandus и евразийския горски подвид Rangifer tarandus fennicus. Евразийския рефугиум най-вероятно е бил малък изолиран рефугиум в Западна Евразия и също дава начало на R. t.tarandus и R. t. Fennicus, което предполага дифилетичен произход на тези два подвида. Южния рефугиум в Северна Америка, простиращ се от Ню Джърси, Кентъки, Мисури, Илинойс и Айова до планинския регион на югозапад – Ню Мексико и Невада дава начало на американското горско карибу Rangifer tarandus caribou. Доказателства за студени, тундрови условия и фосили от тундрови животни, включително карибу са намерени в този регион, определен като Южен рефугиум. По последни публикации, този рефугиум е разделен на три отделни рефугиума, които са се обособили при последното заледяване. На основа на генетични изследвания / секвениране и сравняване на част от контролния регион на цитВ гена от мт ДНК/ на няколко съвременни популации и фосили на Rangifer tarandus caribou , е установено, че съвременните популации се разделят на три хаплогрупи, произлезли от трите отделни рефугиума по време на заледяванията и смесили се бързо при експанзията на вида. Продължава изясняването на точния модел на свиване и разширяване на ареала на вида, от гледна точка, че за видове с начин на живот, адаптиран към студени условия, разширяват ареала си по време на самите заледявания и го свиват по време на затопляне. Следващи изследвания върху филогеографията на северния елен биха дали по-голяма яснота за модела на разпространение на арктични и субарктични животни, което да се има предвид за консервация на вида.
Диви северни елени в Норвегия наброяват приблизително 30 000 и около 10 000 в Свалбард. Популацията в Норвегия се счита за стабилна и се контролира ловуването. Във Финландия горския северен елен R. t. Fennicus е доведен почти до изчезване до 1900 г, но сега възстановява популацията си в резултат на миграция на елени от Карелия и развъждане на животни, които са били уловени и след това пускани на свобода. Горския северен елен остава много рядък за Финландия/ около 1200 индивида в източната субпопулация и около 1000 индивида в западната/. В Европейска Русия северните елени намаляват и вече не се срещат в големи части от тундрата и тайгата. Не е изяснено малката съществуваща популация на Колския полуостров дали е автохтонна или е произлязла от полуодомашнени елени. Подвида R. t. pearsoni обитаващ Нова Земя има малка популация, наброяваща под 1000 индивида, която продължава да намалява. В Азия северните елени са значително повече, отколкото в Европа, макар да няма точни данни за броя им. През 1950 са наброени приблизително 400 000 индивида. В Северна Америка карибу има многочислена популация. В Исландия съществува малка популация от приблизително 1000 индивида, а в Лапландия приблизително 0,5 млн индивида.Стадата на Поркюпайн карибу в североизточна Аляска и прилежащата северозападна Канада, така също стадата от централна Арктика са потенциално застрашени от проучванията и добива на нефт, индустриалното развитие на областите. В Руската федерация най-сериозната заплаха за елените остава бракониерството. Като цяло видът Rangifer tarandus според червената книга на IUCN се счита за слабо засегнат. В Русия видът е включен в червената книга като рядък вид, на който популацията се възстановява. Rangifer tarandus е включен в Приложение III на Бернската конвенция. Подвид R. t. fennicus е строго защитен от Анекс II на Европейската директива за хабитатите и видовете. В източната част на Русия има строги мерки за лов на елени, но въпреки това бракониерството продължава. В Норвегия ловуването също е под строг контрол. Във Финландия строго се ограничава движението на дивите и опитомените стада, за да не се смесят. Вземат се мерки за увеличаване на популацията на R. t. pearsoni на Нова Земля. В Китай видът е рядък, само няколко стотин индивида са останали там. Повечето техни хабитати са унищожени при пожар през 1986г . Включен е в Китайската червена книга. По последни данни като цяло видът има намаляващи числеността си популации /в Британска Колумбия с 40%, в Онтарио 50%, Алберта 60%/, като това трябва да се има предвид за бъдещи мерки. Все повече видът е уязвим от дейността на хората, замърсяванията и от климатичните промени. Ако искаме този вид да не остане само в приказките за дядо Коледа и да се сещаме за него само на Коледа, трябва да вземем необходимите мерки за неговото опазване.
Интересни факти за северния елен
Когато хората мислят за северния елен, обикновено се сещат за Рудолф и неговите приятели, които теглят шейната на дядо Коледа. Много още може да се каже за тези очарователни създания, обитаващи севера на Евразия и Северна Америка.
В Европа ерата на северните елени е била преди 16 000 години, когато те са скитали свободно на стада. Първобитните хора са разчитали на тях за кожи, месо и кости за инструменти.
При северния елен и мъжките и женските имат рога. Женските запазват рогата си, докато родят през пролетта, за да могат да се конкурират за храна. На мъжките рогата опадват всяка зима. Единствено при северните елени всяка година прорастват нови рога.
Северните елени ядат лишеи растящи при субарктичен климат, съдържащи вещества, които спомагат за поддържане на телесната температура и съответно живота на елените през ледените условия на зимата.
Най-големия подвид северен елен е Финландския горски северен елен, който достига до 240см дължина от муцуната до върха на опашката. Финландския горски северен елен има по-дълги крака от останалите подвидове елени. Той е адаптиран към живот в гората и е развил по – широки копита, които му помагат да рови снега за лишеи. Рогата му растат сближени, така че да може с лекота да се придвижва между дърветата.
За саамите, коренното население на северна Финландия, северните елени са важни животни, както за културата им, така и за препитанието. Саамите имат приблизително 400 думи за храна, инструменти, други продукти и части, заимствани от северния елен.
Във Финландия еленовъдството е професия на жителите, населяващи района, докато в други части на Скандинавия, еленовъдството е ограничено само за коренното население.
Мъжкия северен елен издава по-силни звуци, отколкото женския, особено през брачния период. Когато издава рев, той издува малка торбичка, намираща се зад кожата на гърлото.
Северните елени имат силно развито обоняние, което им помага да намират лишеи под снега. Обонянието им позволява да надушат лишеи под 60 см снежна покривка.
Северните елени са приспособени към живот в сняг и студ. Телата им реагират на промяната на температурата с необходимата за това реакция. Когато навън стане прекалено студено, притежават способността да намаляват температурата на техните крайници до нива близки до температура на замръзване. Това позволява тялото им да запазва топлината си и да не измръзва. През зимата лицевата козина нараства така, че да покрива муцуната и по този начин да я предпази от снега.
Може да се каже, че северния елен е една силна, енерго-спестяваща бягаща машина. Малко еленче на приблизително 7 кг, може да надбяга човек, а възрастния елен е по принцип по-ефикасен от всеки друг сухоземен бозайник.
Северните елени имат слаб биологичен часовник. Обитаващите Арктика, където през лятото почти изцяло е ден, а през зимата е нощ, елените нямат нашия 24 часов ритъм на ден и нощ, на сън и будуване. Всъщност те придремват няколкократно през денонощието по различно време и с различна продължителност.
През зимата очите им от златист цвят се променят в тъмно син. Това е приспособление към улавянето на слабата светлина през зимния арктически период.
Наистина северния елен има червен нос като в приказката, най-вече когато е много студено или продължително е дърпал шейна. Това е в резултат на обилното кръвоснабдяване на носа на северните елени с цел да го поддържа топъл, освен това гъсто кръвоснабдения нос участва в регулацията на температурата на тялото на елена.
Козината на северните елени мени цвета си през различните сезони.
За разлика от нас хората, белия дроб на северните елени е триделен. Това се налага поради необходимостта от повече кислород при активните движения на елена.
Имало ли е северни елени на територията на България?
Има няколко факти в подкрепа на това, че този вид е живял и по нашите географски ширини.
– Северните елени представляват една от доминиращите групи животни на така наречената мамутова фауна. В Евразия той се е появил за пръв път през ранния плейстоцен и е доживял до днешен ден. В Източна Европа е отбелязан в средата на плейстоцена. Съвременния ареал обхваща арктическата и субарктическа зона на Стария и Новия свят. В късен плейстоцен е бил широко разпространен в цяла северна Евразия. Максимума на ареала се отбелязва в края на късен плейстоцен. В руските равнини ареала се е съкратил в историческата епоха. Като цяло ареала на разпространение на северния елен е бил свързан със заледяванията и периодите на затопляне между тях, с появата и разтопяването на ледниците. По време на междуледниковите затопляния, северния елен е отстъпвал на север. Екологичната пластичност на този вид му е позволила да се съхрани до сега. Съкращаването на ареала му в днешно време се дължи най-вече на дейността на човека.
– северния елен е мигриращ вид, като такъв извършва големи миграции и се придвижва свободно, с цел намиране на храна и благоприятни условия за живот
– по последни данни кости от северни елени са намирани в Германия, Франция и по най-нови в Полша.
– северния елен плува много добре, т.е реки и езера не представляват географска преграда за него
– Името rangifer, което Линей дава на този вид елен е използвано от Албертус Магнус в не неговото De animalibus : „Dicitur Rangyfer quasi ramifer”. Тази дума може би е заимствана от саамите от думата raingo. За името tarandus, което Линей слага като епитет, се среща в Quadrupedum omnium bisulcorum historia fol. 859-863, Cap. 30: De Tarando на Улизе Алдрованди. Тези имена са споменавани още от Аристотел, Теофраст, както и от Плиний Старши в своята „Естествена история”. Явно е, че такова животно е било познато на тези естественици.
– може сега европейските видове да са по седентарни и да не мигрират както своите събратя от Русия и Америка, но няма доказателства да се счита че са били седентарни и в миналото.
– По данни на Боев З. в публикация за природната среда на България през плиоцена и кватернера, се споменава, че има находки от вида намирани в находища в България, но данните за това не са публикувани.
По следващи изследвания, биха показали до каква степен северния елен е бил разпространен по нашите земи.
Литература и източници.
1. Teresa Piskorska, Krzysztof Stefaniak . Middle Palaeolithic remains of reindeer (Rangifertarandus Linnaeus, 1758) from Biśnik Cave and other cave localities from Poland. Original Research Article. Quaternary International, In Press, Corrected Proof, Available online 16 October 2013
2. Боев З. 2010. Природната среда в България през плиоцена и кватернера – кратък преглед – В.Петков/отг.ред/ България и българите в Европа. Съюз на учените в България. Клон Велико Търново. Изд.”Фабер”, Велико Търново, 366-384стр
3. Cornelya F. C. Klu¨ tsch, Micheline Manseau, Paul J. Wilson. Phylogeographical Analysis of mtDNA Data Indicates Postglacial Expansion from Multiple Glacial Refugia in Woodland Caribou (Rangifer tarandus caribou). PLOS ONE. December 2012 | Volume 7 | Issue 12 | e52661
4. ØYSTEIN FLAGSTAD, AND KNUT H. RøED. REFUGIAL ORIGINS OF REINDEER (RANGIFER TARANDUS L.) INFERRED FROM MITOCHONDRIAL DNA SEQUENCES. Evolution, 57(3), 2003, pp. 658-670
5. Jim Provan1 and K.D. Bennett. Phylogeographic insights into cryptic glacial refugia. Trends in Ecology and Evolution Vol.23 No.10
6. S.A. Byun, B.F. Koop, and T.E. Reimchen. Evolution of the Dawson caribou (Rangifer tarandus dawsoni). Can. J. Zool. 80: 956-960 (2002)
7. Thijs van Kolfschoten a,*, Inge van der Jagt a, Zoë Beeren a, Vasiliki Argiti a, Judith van der Leije a,
Hans van Essen a, Freek S. Busschers b, Pieter Stoel c, Hans van der Plicht. A remarkable collection of Late Pleistocene reindeer (Rangifer tarandus) remains
from Woerden (The Netherlands). Quaternary International 238 (2011) 4e11
8. M. Festa-Bianchet, J.C. Ray, S. Boutin, S.D. Côté, and A. Gunn. Conservation of caribou (Rangifer tarandus) in Canada: an uncertain future. Can. J. Zool. 89: 419-434 (2011)
9. MATTHEW A. CRONIN, MICHAEL D. MACNEIL, AND JOHN C. PATTON. Mitochondrial DNA and Microsatellite DNA Variation in Domestic Reindeer (Rangifer tarandus tarandus) and Relationships with Wild Caribou (Rangifer tarandus granti, Rangifer tarandus groenlandicus, and Rangifer tarandus caribou). Journal of Heredity 2006:97(5)
Този сайт използва ‘бисквитки’ (cookies), за да ви предостави възможно най-добро потребителско изживяване. Можете да промените настройките си за бисквитки, или в противен случай приемаме, че сте съгласни с нашите условия за ползване.ПриемамПрочети повече
Правила на поверителност
Privacy Overview
This website uses cookies to improve your experience while you navigate through the website. Out of these cookies, the cookies that are categorized as necessary are stored on your browser as they are essential for the working of basic functionalities of the website. We also use third-party cookies that help us analyze and understand how you use this website. These cookies will be stored in your browser only with your consent. You also have the option to opt-out of these cookies. But opting out of some of these cookies may have an effect on your browsing experience.
Necessary cookies are absolutely essential for the website to function properly. This category only includes cookies that ensures basic functionalities and security features of the website. These cookies do not store any personal information.
Any cookies that may not be particularly necessary for the website to function and is used specifically to collect user personal data via analytics, ads, other embedded contents are termed as non-necessary cookies. It is mandatory to procure user consent prior to running these cookies on your website.

















 Церес
Церес Bеста
Bеста Ерос
Ерос

 2005UT12
2005UT12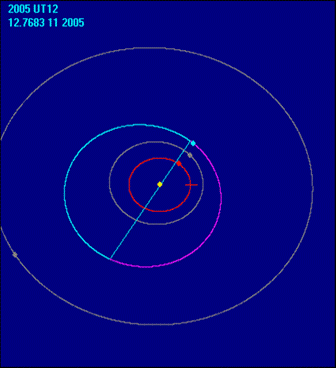 Орбита на 2005UT12
Орбита на 2005UT12 2006ОК14
2006ОК14 2006RJ
2006RJ

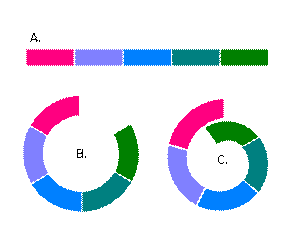

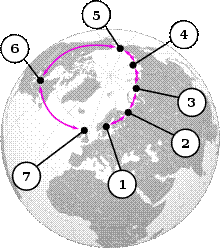
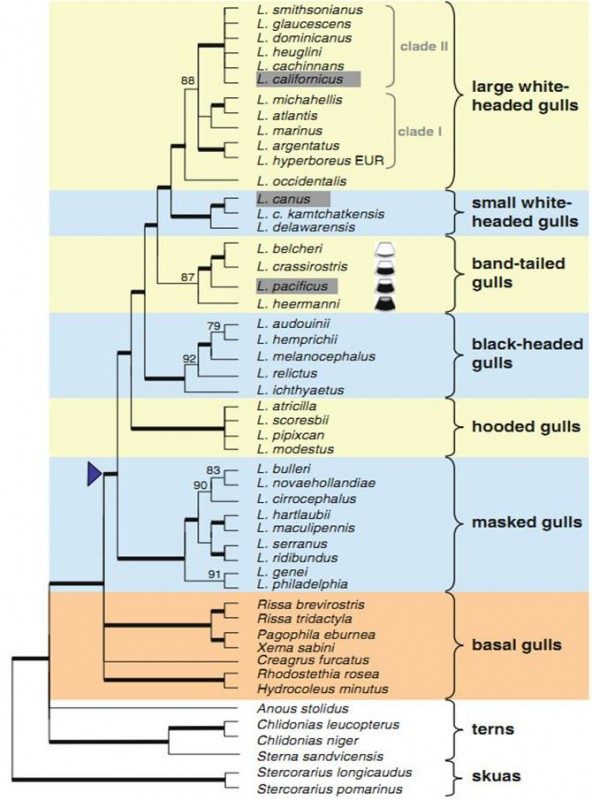

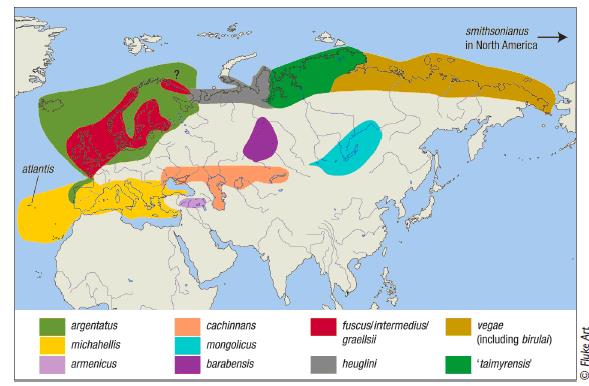
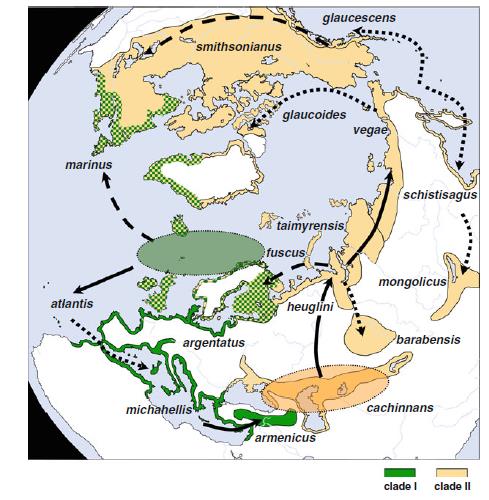 Фиг 2. Филогеография на херинговите чайки – генетично обособени / на основа на мтДНК хаплотипове/ в два клона – Северно Атлантически и Арало Каспийски.
Фиг 2. Филогеография на херинговите чайки – генетично обособени / на основа на мтДНК хаплотипове/ в два клона – Северно Атлантически и Арало Каспийски.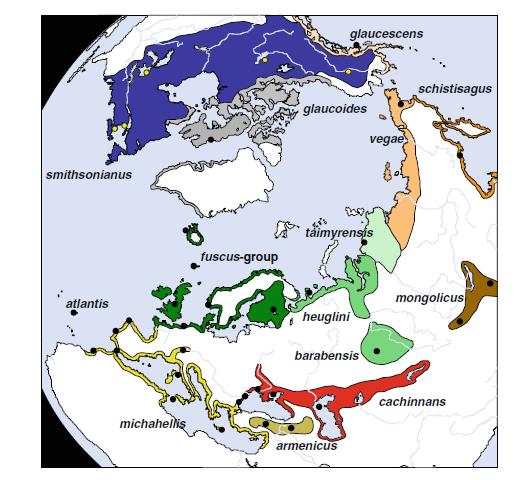
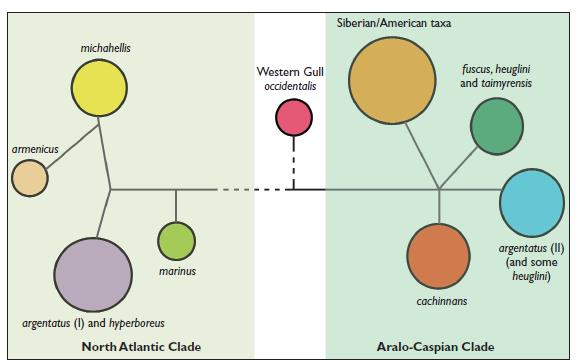

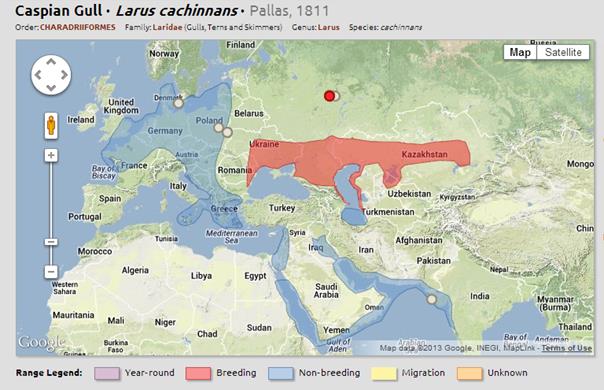


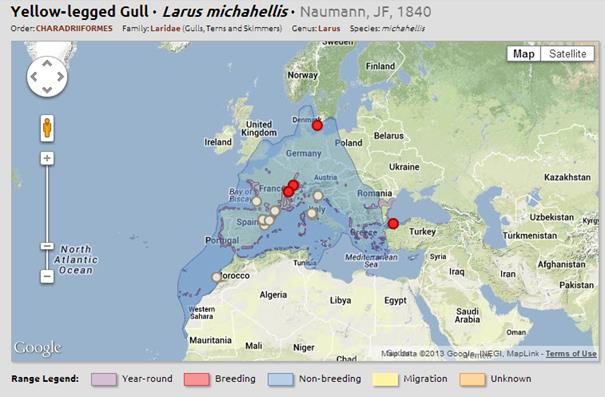







 Дължина на тялото: 41см; Размах на крилете: 120см.
Дължина на тялото: 41см; Размах на крилете: 120см.
 Дължина на тялото: 55см; Размах на крилете: 130см.
Дължина на тялото: 55см; Размах на крилете: 130см.
 Дължина на тялото: 71см; Размах на крилете: 157см.
Дължина на тялото: 71см; Размах на крилете: 157см. Дължина на тялото: 56см; Размах на крилете: 146см.
Дължина на тялото: 56см; Размах на крилете: 146см. Дължина на тялото: 62см; Размах на крилете: 132см.
Дължина на тялото: 62см; Размах на крилете: 132см.



 Северен елен, карибу, северный олень , reindeer, caribou – Rangifer tarandus, Linnaeus, 1758
Северен елен, карибу, северный олень , reindeer, caribou – Rangifer tarandus, Linnaeus, 1758
 Последните елени карибу на Даусон, убити от ловци през 1908 на островите Хаида Гуай. Снимката е след отстреля на животните.
Последните елени карибу на Даусон, убити от ловци през 1908 на островите Хаида Гуай. Снимката е след отстреля на животните.







