Използвайки телескопа на НАСА Спицър, учени са открили кафяво джудже, което е 50 пъти по-масивно от Юпитер, обикалящо с планета около подобна на Слънцето звезда. Такава слънчева система не е била наблюдавана преди, но според учените подобни системи може да са често срещано явление.
Същият екип учени е открил и второ кафяво джудже, което е само 20 пъти по-масивно от Юпитер. Кафявите Т-джуджета са най-студените звезди. А тази звезда е най-младото Т-джудже, което е било откривано някога. Освен това екипа открил и червено джудже, което не обикаля около друга звезда, а се носи само в пространството.Кафявите джуджета са малки звезди, чиято маса не е достатъчна, за да започне ядрено горене на водород в ядрото им. В резултат на това повърхностната им температура е само 1-2 хиляди градуса, а когато остареят температурата им пада до тази на планетите. За това те са тъмни и много трудни за откриване. Първата такава звезда е била открита преди 10 години.
По-масивното от двете новооткрити джуджета се нарича HD 3651 B и се намира в съзвездието Риби. Обекта се намира в слънчева система, чието слънце е по-малко от нашето и бива обикаляно от доста по-малка от Сатурн планета. Орбитата на планетата е силно елиптична, което караше учените да мислят, че някой друг по-масивен обект придърпва планетата с гравитацията си навън. Това със сигурност е Т-джудже. Това е доказателство, че планетарни слънчеви системи могат да приютяват кафяви джуджета, които да правят орбитите на планетите доста странни.
Орбитата на планетата е подобна на тази на Меркурий, но кафявото джудже обикаля на 10 пъти по-голямо разстояние в сравнение с разстоянието от Плутон до нашето Слънце.
Другото Т-джудже се нарича HN Peg B и е от съзвездието Пегас. Докато повечето кафяви джуджета са на милиарди години, HN Peg B е само на 300 милиона години. Учените определили възрастта му като внимателно изследвали акомпаниращата му звезда. Те се формирали заедно от един газов облак. В системата също има диск от прах и скали, който е бил открит по-рано.
Наполеон Бонапарт е роден на 15 Август 1769 година в Корсика. Известно е че той се е родил толкова бързо, че майка му Летиция Рамолино нямала време дори да легне на легло. Той бил втория син на Карло Бонапарт и Летиция Рамолино и бил кръстен с необичайното име ”Наполеон”, заради рогозката, на която бил роден. Семейство Бонапарт били аристократи от по-нисша класа и не толкова богати.
Няколко месеца преди това Корсика бил прехвърлен на Франция от Генуа, така че младия Наполеон се сдобил с нова и могъща родина. Армията го привличала дори и като малък, тъй като тя била една от професиите, за които не била необходима специална подкрепа, която младият Наполеон имал. След като завършил началното училище за момчета в родния си град през Януари 1779г. той бил изпратен с по-големия си брат Джоузеф в колежа в Аутун, херцогство Бургундия. Неговият баща успял да го запише във военната академия в Бриен, от където през Май Наполеон бил прехвърлен в по-модерната военната академия в Париж, докато брат му останал в Бургундия. Военното кралско училище във Франция било най-доброто в цяла Европа преди pеволюцията. Той се дипломирал като артилерийски офицер на 28 Октомври 1785 година, с което започнала и неговата кариера.
За съжаление обаче времената не били добри за Франция. Великата революция тъкмо започвала и в страната бил настъпил смут и бъркотия.
Наполеон бил млад и идеалистичен, поради което се върнал в Корсика да брани независимостта на своята малка родина. Скоро обаче той обърнал гръб на родното си място и се отдал на Франция. Това станало след като участвал в неуспешна атака в Сардиния.
През 1793 година Нополеон постигнал първият си истински военен успех. Това се случило в Тулон, който бил окупиран от англичаните, защото гражданите били монархисти и ги посрешнали без съпротива в средата на революцията. Това разбира се не може да бъде позволено. Идеята на Наполеон била да използва артилерията да си възвърне Тулон. Другите военни лидери също одобрили директната атака. С майсторско артилерийско нападение Наполеон превзел едно от най-важните укрепления и по такъв начин притиснал британския флот. Тулон бил превзет, а Наполеон повишен в бригаден генерал.
През зимата на 1794-1795 Наполеон бил нает в защитата на средиземноморския бряг. Тогава през Април 1795 той бил повикан в Париж и през Юни бил назначен в армията на запада. Той отказал този пост, оплаквайки се от проблеми със здравето. Този отказ почти довел до края на неговата военна кариера, но бил назначен в комитета за национална сигурност.
Женитбата и италианската кампания
За да се докаже отново той искал да бъде изпратен в Турция да сформира султанската артилерия, но съдбата имала други планове. Имало бунт срещу новото управление, който Наполеон трябвало да потуши. Той желаел да се ожени, а вдовицата на генерал Бехарнайс била идеалната партия. Джосефин овдовяла, защото повечето от републиканските привърженици били убити. Нейния съпруг бил екзекутиран, а сабята му била конфискувана. Синът на генерала отишъл при Наполеон да поиска сабята обратно, а той я дал с готовност. Джосефин дошла при него да му благодари и така започнала тяхната връзка. Представител на аристократическата класа тя видяла в младия генерал възможност да бъде защитена от революционната бъркотия. Сватбата се състояла на 9 Март 1796г. Била малка церемония без църква, но в присъствие на магистрат. Наполеон дори закъснял с 2 часа, което говори много колко истинска била неговата любов. На 11 Март меденият месец трябвало да приключи, защото той трябвало да замине за Италия, назначен за командир на италианската армия. Безполезно е да се каже, че Джосефин не е била сама, докато го е нямало. Армията, която той трябвало да ръководи не била в добро състояние, както и останалите републикански армии. Нямало достатъчно пари и храна. Тя била снабдена със 150 коня и магарета, който също не били достатъчни за тогава. Цnbsp;ъководителите били крадци и прибирали всичко, което можели и живеели по крайбрежните градове. Генералът, който Наполеон трябвало да смени бил 60 годишна мумия, който нямал ни най-малка представа как стоят нещата извън неговия офис. Новият генерал трябвало да промени доста неща. Той трябвало да формирова войските отново, които имали нужда от повече морал и цел към, която да се стремят. Той взел 7 милиона лири заем, който бил използван за поддържането и подобряването на боеспособността армията. Тези бедни войници, които Наполеон трябвало да води, нямали нищо и биха последвали всеки, който им обещае богатство. А той направил точно това и знаел как да осъществи обещанията си. След като последвали Наполеон в битка тези мъже станали нещо повече, а имено наполеонави войници, гордеещи се с това и служещи му напълно верни. Той се грижел за своите войници, например след въстанието в Милано той им позволил за три часа да разграбят града. Това било сторено без пощада, като войниците останали доволни. По време на война с Австро-унгарската империя, в която Наполеон участвал като командир на трета армия на страната на Франция. Неговата мисия била да превземе долината По, което не била лесна задача, тъй като враговете били навсякъде. Той се интересувал и спазвал заповедите на управляващите, което го правило просто брилянтен и успял да осъществи мисията си напълно. Също така и грешките на противника му дали голамо предимство. Наполеон винаги се стремял да има числено превъзходство, което той постигал като разделял вражеските сили на малки части чрез бързи движения. Теренът бил много труден, мостовете и реката изиграли важна роля в тази война. Първо той разбил Пиедмонт на 14 Април 1796 при Милесимо и го изкарал от войната. На 15 Май Наполеон насочил войската си към Милано, след което към владенията на Парма и Модена да възстанови мира. Всяка победа била заплащана с цената на много жертви. За да защити стабилността на всяко свое завоевание, той сформирал две републики, които взаимствали своята конституция от Франция. Това било велика стъпка, показваща, че Наполеон разбира не само от военни дела, че е повече от войник и че има потенциала да бъде велик човек. На 8 Октомври той обвинил Модена, че прави заговор с Австро-унгария. На другия ден генералът дал своето съгласие на Генуа да стане протекторат на Франция. На 10 Октомври Наполеон подписва мирен договор с Неапол. Където и да отиде той установява мир и ред. Най-важните битки от този военен поход били битката за моста Лоди на река Адда на 10 Май и битките на 16 и 17 Ноември на моста Аркол. Той отново трябвало да окуражава своите войници, тъй като всички били уморени от войната. На 14 Февруари Наполеон победил при Цnbsp;ива и при Таглименто на 16. Той навлязъл дълбоко в територията на врага като се насочил към Виена. Било сключено временно примирие на 18 Април 1797г. в Лебен, а по-късно на 18 Октомври 1797 и мирния договор Кампоформио бил подписан. Мирът бил сключен за период определен от Наполеон, а не от правителството. Той вече бил победител, а известността му нараснала.
Египетската кампания
Нови мисли засягащи изтока вълнували съзнанието на Бонапарт. Френските управници били доволни да се отърват от известния генерал и одобрили неговите планове да установи френско влияние в Египет, с цел да откъсне Индия от Великобритания. На 19 Май 1798г. Наполеон отпътувал от Тулон с армия от 35 000 човека. На 11-12 Юни той завладял Малта, а на 30 вече бил в Александрия с цялата си войска. Градът бил превзет, а той се отправил към Кайро по западния приток на Нил. Първата битка се състояла на 12 Юли в близост до града и пирамидите. С минимални загуби Французите отблъснали врага и Египет паднал под контрола на Наполеон. Той променил правителството, пощата и системата за събиране на данъци, въвел печатната преса, направил министерство на здравеопазването, построил нови болници за бедните в Кайро и открил Египетски институт. Но през този период британците не бездействали. На 2 и 3 Август адмирал Нелсън атакувал и унищожил френската флота в Абокирският залив, оставяйки френската армия без връзка с Франция. Турският султан започнал нова офанзива в Египет, а по същото време Наполеон претърпял загуба при атаката си на Сирия. Върлуващата чума убила много хора от войските му и дисциплината се разваляла. Той имал една ненужна победа в Абокир на 25 Юли 1799г. срещу турците. Наполеон поверил командването на генерал Жан Колбер и се върнал обратно във Франция с няколко офицера. Това било една голяма погрешна стъпка от страна на Бонапарт, която се превърнала в приключение с цената на много човешки животи без никаква изгода.
Консулското звание
На 9 Октомври 1799г. Наполеон акустирал и се отправил директно към Париж. Обстановката там била подходяща за големи промени. След Египет Наполеон започнал да се придвижва на политическата сцена, разчитайки на подкрепата на хората и което е по-важно армията стояла зад него, въпреки неуспеха в Египет. Цnbsp;ечтта, която Наполеон държал пред Барас била близка до чувствата на хората. От нея става ясно, че той вече не служи на никого освен на народа си. Той също казал, че без него Франция нямало да съществува, а хората, които управлявали я превърнали в руина и изтъкнал, че има желание да спаси страната си отново. Хората се съгласили, че Франция е готова за диктатор след ужасите на управляващите и прочистванията. Пет от членовете на управлението били наричани ”петте маймуни”. Животът на хората не се е променил в равенство между всички. Също се носели слухове за английски заговор. Наполеон бил назначен за главнокомандващ на военните сили на Франция. На 9 Декември 1799г. започнал със своите промени. Отначало всичко вървяло добре, но тогава той срещнал затруднения със съвета на петстотинте и когато влязъл в залата бил наречен ”изгнаник”. Луциен Бонапарт, неговия полу-брат, който бил глава на съвета, бил задължен да извика войниците да помогнат. Тогава влязъл генерал Леклер, водач на пехотната гвардия и помогнал на Наполеон. Членовете на съвета започнали да бягат във всички посоки, дори и през прозорците, оставяйки скъпите си якета и връхни дрехи в залата. Така успехът бил постигнат, а републиката фактически свършена.
На 13 Декември новата конституция била приета, в която Наполеон легализирал своите реформи и действия. Франция е сменяла конституцията си много пъти до тогава по време на революцията, а тази била почти като другите, но това имало малко значение. Сега имало три консула, от които Наполеон бил първи и най-важен, другите били просто играчки за проявите. На 19 Февруари 1800г. той се преместил в Тюлер, където Наполеон било добре дошла изненада за много. Той работил усърдно и се опитвал да узнае всичко за всички. Той систематизирал закона, стабилизирал валутата, поправил закона за данъците, подобрил образователната система, успокоил религиозните въпроси в страната и т.н. фактически може да се каже, че той е създал съвременната администрация. Също така установил примирие с Папата на 15 Август 1800г. Сенатът го обявил за пожизнен консул на 2 Август, когато мирният договор с Великобритания бил обявен.
Преди този мир обаче имало още една война. Наполеон предложил примирие на Австрия и Англия на бъдни вечер 1799г., от което те не били заинтерисовани. Трима френски посредници са убити в гората близо до Цnbsp;адщад. Пролетта на 1800 донесла война в Европа. Френската армия от Цnbsp;ейн под командването на Морьо надвили австрийците и ги отпратили отвъд Дунав. Цnbsp;уснаците също застанали срещу Франция. Страхливият генерал Суворов ударил Французите в Италия два пъти. Наполеон напуснал Париж на 6 Май и се насочил към Алпите. За пет дена от 15 до 20 Май той прекосил клисурата Св.Берханд. на 2 Юни Бонапарт достигнал Италия. Неговият отличен план бил да заблуди врага да постави главните си сили пред него. Той планирал като едно от главните си задължения да прекрати тази война и успял. На 14 Юни 1800г. в Маренго се състояла битка. Някои мислят, че тази битка е моментът, в който била създадена Френската империя. Всъщност Маренго били три различни битки близо до Александрия. В шест сутринта френските войски се появили на полето. Две френски укрепления издържали 4 часа, но са били принудени да ги напуснат. Бонапарт не може да преодолеят австрийците, а положението става все по лошо за него. Спасителят този ден бил Десае, който пристигнал точно навреме и французите взели превес. Австрия била победена, а войските й изпратени в Бормида. Наполеон изпратил на австрийския император писмо, в което му предложил да се предаде. Писмото било написано като между равнопоставени личности, Наполеон не се смятал за консул или генерал. Но мирът не дошъл от първия път. След Маренго имало истински тържествен дух във Франция, но той бил твърде ранен. Преговорите в Лунвил през Ноември 1800г. били в застой. Военните действия се възобновили на 22. при Хоенлинд генерал Морьо донесъл на Франция голяма победа над австрийците на 3 Декември 1800г. австрийският херцог Чарлз загубил 12 000 мъже за две седмици от сраженията и 25 000 заложници от 70 000 армия. Морьо достигнал на 75 километра от Виена и притискал Австрия да приеме мира. Генералът бил главния наполеонов съперник особено след тази битка. По-късно той участвал в заговор и бил заточен в Америка. Той стоял там от 1804 до 1813. след като се върнал в Европа той участвал в сражението за Дрезден на 26-27 1813г срещу Наполеон и бил убит. Мирът бързо се разпространил след като и руснаците излязли от войната. Великобритания останала сама подписала горепосочения мир. Франция също била в мир и могъща. Много републики изникнали около нейните граници, всички под ботуша на Наполеон. Например република Цизалпик била създадена в Лион, Франция, а не в Италия. Докато нямало способни италиански представители, царуването било предадено на Наполеон. Австрия изгубила всичките си италиански области, но получила Венеция, която след векове република престанала да бъде такава.
Бонапарт бил пожизнения консул на Франция, но това не било достатъчно. Унаследената монархия била финала. Много опити за убийство срещу Наполеон направило решението му лесно. Добър пример за тези опити бил през Декември 1800г., когато бомба убива 22 и ранява 57 човека и разрушава 42 къщи. Наполеон преминал през мястото малко по-рано. Сред осъдените за този инцидент бил художникът Керачи, който бил приятел от дедството на Наполеон. Друга известна и съдбоносна конспирация срещу Наполеон била водена от генерал Пичегру и Джордж Кодудалин. Това дало основание да заловят младият херцог на Енгиен, който бил консерватор. Френските войници го заловили в друга страна и го закарали във Франция да бъде съден. Никакво доказателство не било намерено, но въпреки това херцогът бил екзекутиран. Това довело протести на омраза и неодобрение от цяла Европа.
Империята
До 1802 г. Наполеон се превърнал в най-известният диктатор, който Франция е имала някога. На 18 Юни 1804г. сенатът обявил, че първият консул е императорът на Франция, с божие благоволение и според републиканския закон. В Нотр Дам на 2 Декември Наполеон получил своята корона. Лично папата присъствал, така че възкачването на новия император да бъде законно сред неговите равни в Европа. Политическата обстановка станала любопитна. Въжможен ли е мир между европейските нации, които са се борили срещу революционна Франция и новата Франция, която била вече монархия базирана на революция? Той направил чудесен дворец за себе си, пищност и слава, величествена перспектива за хората. Били раздадени и благороднически титли като например ”гранд адмирал” и др. Нищо от това не можело да предотврати бъдещите войни.
Победи по целия континент
През 1805г. бил създаден тристранен съюз между Англия, Австрия и Русия, в който участвала и Прусия. Те обединили силите си, заради мирните договори от 1802 и 1803г., където Франция е взела твърде много за себе си. Наполеон поел властта на Щвейцаря, станал президент на Италия, много италиански области били присъединени към Франция като Генуа, Неапол и пристанищата. Германия била напълно пренаредена, някои бивши австрийски области били предадени на Прусия, Баерн и Вуртемберг. Наполеон не чакал и изоставил своите планове за нападение на Англия, така че бил свободен да започне война. Той направил Великата Армия. Войсковите части се състояли от областни войски, поради което трудно се справяли с ръководенето. В този момент той съсредоточил армията си в Болоназа нашествието в Англия, но това не се случило и вйската тръгнала срещу Австрия. В Улм те били разбити от французите. След като маршал Ней пресякъл пътя на оттеглящите се австрийски войски над 50 000 австрийци били пленени на 20 Октомври 1805г. ден след като френската флота била разбита от Хроцио Нелсън при Трафалгар. Френският адмирал Вилньов против желанието му бил изпратен в Средиземно море от Наполеон. Адмиралът загубил при Абукир. Френско-испанската флота изгубила 18 кораба от 33. Малка утеха за Наполеон била новината, че Нелсън е умрял, застрелян с пушка. Той държал британските морски владения, което осигурявало сигурността на Англия. Двете страни се срещнали в Аущерлиц, което е малко село днес. Преди това, на маршал Ней били дадени заповеди да отблъсне австрийците извън Тирол. Той също трябвало да завладее Виена. Бонапарт продължавал напред, докато накрая не се сблъскал с битката, която търсел. Той избрал мястото като завел врага, където искал. Австрийците и руснаците се опитвали да го спрат от завръщането му във Виена. На 29 Ноември Наполеон напуснал Аущерлиц да се срещне с руско-австрийските войски под командването на Александър II и Франс II, които се опитали да обградят французите. Битката при Аущерлиц е може би една от най-добрите на Наполеон. Той бил готов на всичко, което врага опитал и отвръщал на всичко със защита. На сутринта на 2 Декември започнало самото сражение. В 4 часа следобед всичко приключило. Австрийците и руснаците загубили 35 000 мъже, 50 флага и цялата си артилерия., тогава тя била стопена за паметник на победата и откарана в Париж. Френският император загубил 8 000 мъже. На 6 Наполеон се срещнал с Франс II, но не се стигнало до мир, въпреки споразумението от Пресбърг от 26 Декември 1805г., където Австрия загубила Венеция. Така Австрия напуснала четворната коалиция. Наполеон първо трябвало да изведе Прусия извън войната, а тази в Саксония нямала толкова голямо значение за него и на 28 Октомври 1806 той превзел Берлин. В битката за Ейла на 2 Февруари 1807г. Наполеон бил победен от руснаците, а цената била огромна. Тази война продържила 6 месеца и на 25 Юни Александър и Наполеон се срещнали в Тилист. Победителка била Франция, а Цnbsp;усия все още била неукротима. След тази среща всичко изглеждало добре за Наполеон. Той получил правото да преоформя западна и централна Европа както желае. Най-значиммият резултат бил създаването на херцогство Варшава през 1807г. Също Швеция била победена с руска помощ на следващата година. Така Франция се превърнала в господар на целия континент. Англия все още не била засегната. Тя била по-силна от всякога и сякаш никой не можел да й се противопостави. Но императорът направил грешка.
Проблеми с Испания и Англия
През 1808 Наполеон изтласкъл бурбоните от Испания и поставил брат си Джозеф на трона. Испанците обаче не го харесвали и мразели Франция, защото крадяла пари от църквата, а и не искали да бъдат контролирани от тях. Войната била неизбежна. Великата Армия сега се изправила пред враг, който бил мотивиран от патриотизма религията и било невъзможно да бъде притиснат. Гуерилската война била нещо, с което великата армия не могла да се справи. В Байлен на 23 Юли 1808г., 18 000 френски войници трябвало да се предадат. Това било шок за цяла Европа. Испания накрая подчинила 300 000 мъже от великата армия от войната на полуострова. Тази война също била съдбоносна, защото Англия вече можела да започне нейната война по суша. На 1 Август 1808г., 15 000 мъже под командването на Артур Уелъсли, херцогът на Уелингтън слязъл на португалския бряг. Всичко продължило 6 години до 1814 в Тулон. Във Витория Уелингтън спечелил най-голямата си победа в Испания. За сега Наполеон си мислил, че е укротил испанците след превземането на Мадрид на 5 Декември 1808г., но грешал. Австрийците отново били готови за война и през 1809г. тя започнала. Един млад австрийски студент се опитал да убие Наполеон в Шонбрън с кама, а и имало голямо неспокойствие в германоговорящите области. Между 19 23 Април, за 5 дни, Наполеон покосил и разпръснал Австрийците. Но този път той не победил толкова лесно. Когато се опитвал да прекоси р.Дунав в Асперн и Еслинг на 22 Май бил победен, а войските му изпаднали в ужасно затруднение на остров Лобау. Австрийците били командвани от херцог Чарлз, който бил един от най-упоритите опоненти на Напоеон. Загубите в двете страни били големи. Там Наполеон загубил маршал Лане. Двудневната битка в Ваграм донесло на Бонапарт победа на 5 и 6 Юли. Тази война била близо до укриващият се Наполеон и дори Англия стигнали остров Валхерн в Холандия, но отстъпили. Австрия сключила мир с Франция във Виена на 14 Октомври.С настъпването на мира Джосефин се развела с Наполеон през Март 1810г. той сключил брак с херцогиня Мария Луйза, дъщерята на Франс II. Този брак задържал за известно време мирът между двете нации. Наполеоновият син се родил на 20 Март 1811г. брат му Луйс, крал на Холандия; Джером, крал на Уестфален; Джосеф, крал на Испания и неговият полубрат Луциен вече не били най-близките му, той имал наследник. Войната с Цnbsp;усия била все още на хоризонта. Наполеон опитвал много пъти да сключи мир, но без успех. Конференцията в Ерфурт между 27 Септември и 14 Ноември 1808 не донесла отговари на много важни въпроси. Така Франция и Великата Армия били въвлечени отново във война с Цnbsp;усия. 700 000 мъже щели да отидат там, от които 300 000 в действителност французи. Нямало връщане назад, тъй като самият Наполеон се насочил натам. Цnbsp;уснаците смятали да се защитават от французите като построят крепостни окопи, където наполеоновите войски ще загинат под огъня на оръдията. Обаче разположението на тези окопи било избрано неточно и Наполеон трябвало да залови руската армия без да отстъпва към капана. Така той се предвижвал, но руснаците осъзнали грешката си точно навреме и успяли да се имъкнат от техния собствен капан. Наполеон разбрал, че ако започне да преследва вражеските войски из цяла Цnbsp;усия ще бъде гибелно за неговата армия. Но ако се върнел обратно в Европа това щяло да означава, че е загубил. Дилемата, пред която бил изправен била дали да преследва руснаците с надеждата за битка, която да спечели и да сключи мир. Войната започнала през 1812г., а Наполеон трябвало да реши този проблем един път за винаги. Неговите войски се отправили към река Днепър и я прекосили на 15 Август. Цnbsp;уснаците напуснали град Смоленск на 17 след кратка съпротива. Когато започнала същинската битка в Бородино на 7 Септември, Наполеон не бил подготвен и руските войници нанесли ужасни поражения на караулите. Победата била за Кутузов, който трябвало да защитава Москва. Наполеон отхвърлил всякакви маневри и заповядал директен фронтален щурм. В края той успял да избута руснаците от техните позиции и нощта паднала. На сутринта руската армия се оттеглила, но битката не била спечелена за Наполеон. Александър дори отказал споразумение с Бонапарт, който завладял Москва, но това с нищо не му помогнало. От тук нямало изход на никаде. Това доказва, че френските войски не били готови за такъв тип военен поход, а и често било невъзможно да се намерят запаси, откакто казаците били отслабили фланговете. Но зимата се оказала най-големия враг от всички. На река Брезина на 25 Ноември 1812г. наполеоновата армия претърпяла тежки поражения главно от времето. Над 350 000 мъже умрели. Императорската охрана намаляла от 30 000 мъже на 1500. Наполеон изоставил своята Велика армия от около 500 000 човека в Цnbsp;усия. Било ясно, че Европа ще се обърне против него. В Париж дори имало сполучлив военен преврат на 23 Октомври воден от генерал Малет, който бил екзекутран. На 23 Юни 1813 принц Метерних се срещнал с френския император в Дрезден. Той отказвал да сключи примирие откакто победил в Лутцен на 1 Май и в Баутцен на 16 Май 1813г. Наполеон вече имал нова и млада армия, наричана още ”армията на Мария Луиза”. Той разполагал с нови 300 000 войници, но качеството не било както преди. Вече не сключвал мир, тъй като вярвал, че може да победи. Не било необходимо да се броят враговета, защото би могло да се каже, че Франция била сама срещу цяла Европа, разчитайки само на поляците. Австрия, Цnbsp;усия, Англия, Прусия всички те били в нов съюз. В Лайпциг на 16-18 Октомври се състояла битката на нациите. 150 000 френски войници се сблъскали с руски, австрийски и пруски войски от около 300 000 човека. Първият ден изглеждал добре за Наполеон, но след това той загубил, сякаш късметът му го напуснал. Франция вече имала репутацията на самотната страна.
След кото изгубил Германия Наполеон се борил вв Франция. Френският военен поход на Бонапарт бил неговия най-добър, истинско доказателство за неговия стратегически гений. Но в края той почти не играел важна роля. Наполеон разполагал само със 70 000 мъже, повечето от които новобранци, за които съюзниците мислели, че ще преборят лесно. Те смятали да превземат Париж за една седмица, но всъщност им отнело три месеца. Бързи маневри и светкавични атаки направили тъжен живота на съюзническите армии. На 11 Февруари 1814г. в Монтмирал и на 18 Февруари в Монтрьо били най-големите успехи на Наполеон. По-късно и прусите били жестоко разбити. 31 Март бил денят, в който врагът достигнал Париж. Маршалите на Наполеон го карали да абдикира. Той го сторил на 4 Април 1814г., притиснат от безнадеждната военна обстановка. Опитал се да направи синът си Наполеон II следващия управник но 3 годишното дете не било признато от съюзниците, а на трона бил възкачен Луй XVIII. Той бил изпратен на заточение на остров Елба, където щял да получава 1 милион франка годишно и да управлява острова. В същото време съюзът се разпаднал по време на конгреса във Виена, а французите не били удовлетворени от възвърналите се монархисти. Наполеон имал планове и след десет месеца на 26 Февруари 1815 той напуснал острова и се отправил към Франция със 1050 войника. На 1 Март пристигнал в Антибес, след което пътувал 20 дни до Париж необезспокояван. На 21 Март Наполеон влязъл в Париж подпомогнат от бурбоните. Лудвиг XVIII бил прогонен и с големи мъки трябвало да унищожи всичко, което било свързано с революция. Бонапарт взел знамето на революцията отнова и се узаконил пред французите, въпреки че те не били убедени. Френският народ не желаел възобновяване на борбата срещу Европа, но Наполеон трябвало да я възобнови ако желаел да остане на трона, въпреки че и той не искал повече война.
Битката при Ватерлоо
Военният поход при Ватерлоо (12-18 Юни) бил кратък и решаващ. По навик първата атака била дело на Наполеон. При Ватерлоо той загубил за последен път. На 16 Юни наполеоновите планове да раздели пруските и английските войски били почти осъществени. В Лигни той разбил прусите, но не успял да вземе превес над тях. Те се прегрупирали и се придвижили към Ватерлоо два дни по-късно. По същото време Ней не успял да превземе кръстопътищата при Куатре Брас, а Уелингтън го държал в залива. В крайна сметка Ней се сблъскал с цялата анклийска войска. Ватерлоо бил на практика най-съществения момент. Наполеон се опитвал цял ден да смаже англичаните, но Уелингтън едва успял да удържи позициите си. Бедата дошла, когато 10 000 кавалеристи без подходяща артилерийска подкрепа и пехота атакували британските редици и се провалили. Следобеда Маршал Ней успял най-накрая да направи добра атака, но прусите разкрили присъствието си по десния фланг, а нямало други резерви, които да подпомогнат Ней. Прусите били отблъснати с помощта на караула, който по-късно се насочва към много отслабналите английски редици, но това била много груба грешка. Те били разположени на лошо място, където английските пушки и мускети нанесли огромни поражения, може би убивайки почти половината от тях. Наполеон се намирал в своя край като понесал последното си поражение на 18 Юни. С остатъка от армията си той поел към Париж, където на 22 Юни абдикира за втори пореден път.
Наполеон се опитал да избяга в Америка, но на 3 Юли 1815г. бил заловен на пристанище след като то било блокирано от английски военни кораби. Той решил да се предаде на британците и помолил за сигурно убежище. Съдбата му била заточение на остров Св. Елена като служител на британският гобернатор сър Хъдсън Лов. Той тръгнал на 8 Август. Животът му там не бил особено приятен, тъй като му липсвал синът му, който се намирал във Виена. Храната не била добра, а климатът нездравословен. Така на 5 Май 1821г. Наполеон Бонапарт починал на 52 годишна възраст след дълго боледуване от рак. Неговите останки сега се намират в църквата ”Les Invalides” заедно с някои от неговите маршали. Близо до църквата на подобаващо място е разположен военният музей, в който са представени много от знамената от времето на неговото управление.
Исак Нютон (1643—1727 г.) — един от най-великите физици на всички времена. Роден е на 4 януари 1643 г, в с. Булстроп, графство Линкълншир. Баща му Исак Нютон—дребен земевладелец, умрял малко преди раждането му. Самият Нютон се родил преждевременно, хилаво и болнаво дете за чийто живот близките му доста се страхували. Въпреки това той живял, без да боледува и доживял до дълбока старост. След завършване на селското училище на дванадесетгодишна възраст Нютон продължил образованието си в градското училище в съседния Грантъм.
Оскъдните сведения, запазили се за ученичеството на Нютон, ни го рисуват като срамежливо, затворено момче, което избягвало шумната компания на своите съученици. Заедно с това се отбелязва и неговото самолюбие — Нютон не понасял обиди и в желанието си да изпъкне сред съучениците си, ако не със сила, поне с успеха си, той се издигнал от посредствен ученик на едно от първите места.
Обучението в грантъмското училище имало схоластичен средновековен характер. Освен богословски предмети — свещено писание, библия и пр., Нютон изучавал още древни езици — латински, гръцки, староеврейски, които овладял основно. Що се отнася до новите езици — френски и немски — тях Нютон владеел слабо. Училището не му дало достатъчни познания и в областта на математиката. Той изучавал само аритметика и елементи от геометрията.
В свободното си време Нютон се отдавал на изработването на механични играчки и уреди, увличал се в изкуството — рисуване, писане на стихове, четял много и с желание.
През 1658 г. петнадесетгодишният юноша Нютон завършил училище. Под влияние на неговия вуйчо Ейскоу, получил образованието си Кеймбридж и на неговия учител Сток, Нютон се готви да постъпи в Кеймбриджския университет. На 5 юли 1661 г. той е записан като студент в Тринити колидж. От този момент до 1696 г. животът на Нютон е свързан с Кеймбридж, с Тринити колидж. Тук той последователно преминава всички степени — от студент до член на колежа (преподавател) и професор. Тук той е създал най-великия си научен труд — „Математически принципи на естествената философия“ (или накратко „Начала“), направил е най-големите си открития.
В Кеймбридж се развил с целия си блясък научният гений на Нютон. Би могло да се помисли, че превръщането на Нютон от тихо и срамежливо момче от село в най-велик световен учен е станало в резултат на влиянието на специфичната научна атмосфера в Кеймбридж, обаче това не е вярно. Тъкмо обратното: Нютон станал Нютон не толкова благодарение на Кеймбридж, колкото въпреки него. Всъщност заедно с растежа на Нютон се развил и Кеймбриджският университет, който се превърнал от изостанало средновековно учебно заведение в най-крупния център на съвременната наука.
1664 г. е превратна година в живота на Нютон. Той преминава в горния курс и получава званието действителен студент. В този период той замисля своите велики открития. През януари 1665 г. Нютон получава степента бакалавър и заминава в родното си село поради чумна епидемия. Тук, на село, той окончателно обмислил своите открития: новия математически метод — смятане с производни, разлагането на светлината и законите за всеобщото привличане.
От 20 май 1665 г. до началото на 1666 г. Нютон написал пет статии за новия математически метод, но нито една от тях не била публикувана. Любопитно е да се отбележи, че Нютон по принцип не обичал да бърза с публикуването на своите открития. Понякога създателят на новото, намирайки се още в плен на старото, сам е пречел за разпространението на новите идеи. С това Нютон правел лоша услуга както на себе си (по-късно той е бил въвлечен в тягостния спор с Лайбниц относно приоритета в откриването на диференциалното смятане), така и на развитието на науката. В 1667 г. Нютон бил избран за младши член на Тринити калидж, а след няколко месеца, на 16 март 1668 г. — за старши член. След година и половина, през октомври 1669 г., неговият учител Бароу му предал своята професорска катедра.
Двадесет и седемгодишният Нютон станал професор в кеймбриджкия университет. Оттогава Кеймбридж започнал да се слави не с богословие, а с физика и математика. Получаването на катедрата, в която работил Нютон, станало въпрос на чест за английските учени.
Какви заслуги е имал младият учен, за да бъде издигнат до такъв висок пост? Както се споменава по-горе, Нютон в продължение на две години, 1665—67 г., по същество вече направил своите велики открития, но това не било известно никому — Нютон не е имал печатни трудове. Въпреки всичко това славата на Нютон се разпространила в Кеймбридж и Бароу, споменава в печата за забележителните способности и знания на Нютон. Без съмнение, владеейки нов, по-могъщ математически метод (диференциалното смятане), Нютон без особени затруднения решавал най-сложните математически задачи, които циркулирали в тогавашния учен свят.
Нютон имал и други твърде осезаеми резултати. Става дума за неговите работи в оптиката, които били главното му занимание през първите години на професорската му дейност (той започнал да чете след Бароу лекции по оптика). В резултат на продължителни проучвания за усъвършенстване на телескопите той конструирал през 1668 г. модел на нов телескоп – рефлектор. Последният бил твърде миниатюрен апарат, дълъг само 15 cm, с диаметър 2,5 m, но давал същото увеличение като двуметрова тръба. След три години през 1671 г., Нютон построява втори телескоп-рефлектор с дължина (около) ~ 120 cm, с радиус на огледалото повече от 2 m. Този телескоп предизвикал голяма сензация Той бил изпратен в Лондон, където бил разгледан от краля и членовете на Кралското дружество (Английската академия на науките). Уредът получил единодушно одобрение. На 11 януари 1672 г. Нютон бил избран за член на Лондонското Кралско дружество.
Нютон не се съмнявал във важността на направените от него научни открития и бил уверен, че те ще бъдат посрещнати от учения свят още по-благосклонно, отколкото например телескопът. Но него го очаквало жестоко разочарование. Новото не побеждава без борба — в тази истина е трябвало да се убеди Нютон след като на 6 февруари 1672 г. той изнася доклад в Кралското дружество върху новата теория на светлината и цветовете. Против опитите и особено против изводите на Нютон заставал членът на Кралското дружество Робърт Хук (1635—1703 г.), знаменитият физик Християн Хюйгенс (1629—1695), йезуитите Пардис, Лилус, белгийският учен Люкас и др. При това доста интересно е обстоятелството, че наред с неразбирането и непризнаването на новото, внесено в науката на светлината, го обвинили в плагиатство, в заимствуване на чужди открития Такова обвинение например повдигнал Хук, приписвайки на себе си приоритета за опитите с оцветяването на тънки пластинки и оспорвал приоритета на Нютон в изобретяването на рефлектора. Такива обвинения в присвояване на приоритет съпътствали всяко откритие на Нютон: приоритета за изобретяване на диференциалното смятане оспорвал Лайбниц, а за откриването на закона за гравитацията — пак Хук. Всички тези неща показват, че откритията на Нютон, при цялата им дълбочина и значение, не са били спонтанен изблик на откровение, а са се подготвяли от цялото развитие на науката, или както се казва „са витаели във въздуха“. Заслугата на Нютон е, че той е довел тези идеи до такова съвършенство и завършеност както никой друг от неговите съвременници. Той по-дълбоко и по-далеч от всички проникнал в новото, което се е заключавало в тези идеи. Затова с пълно основание считаме единствено Нютон за автор на споменатите открития.
Преминал от въпросите на оптиката към въпросите на гравитацията и механиката, Нютон работи над тези проблеми интензивно. По онова време мисълта за силата, която заставя планетите да се движат около Слънцето по законите на Кеплер, вълнувала много умове. За тази сила мислил и Хук, който доловил правилния й закон, а също и членовете на Кралското дружество, известният лондонски архитект Кристофър Рен и астрономът Халей. Те дори успели да формулират съответната задача: да се докаже, че под действието на сила, обратно пропорционална на квадрата на разстоянието до Слънцето, планетата ще се движи по елипса, но не могли да я решат. Халей решил да се обърне към Нютон за помощ. През август 1684 г. той бил в Кеймбридж и се отбил при Нютон, за да се консултира с него по тази задача. Нютон отговорил, че е решил вече задачата и обещал да му изпрати своя ръкопис. През есента ръкописът бил у Халей. Халей незабавно поставил пред дружеството въпроса за публикуването му. Но Нютон не се съгласявал да се отпечата представената част от неговия труд. Едва на 28 април 1686 г. той представил пред Кралското дружество ръкописа на книгата си „Математически принципи на естествената философия“. Отпечатването на книгата се забавило по различни причини, но в края на краищата всички пречки били отстранени и в средата на 1687 г. великото творение на Нютон се появило на бял свят. С издаването на тази книга се завършва епохата на борбата за нова наука и мироглед и започнала нова епоха в развитието на науката.
Преди да дадем представа за общата схема на „Началата“, би могло да отбележим, че в механиката геният на Нютон се проявил преди всичко в осмисляне работите на неговите предшественици и в обобщаване на закони, известни дотогава за частни случаи.
Първите страници от „Началата“ съдържат основните понятия, аксиоматиката на класическата механика. Тази част се състои от осем определения, три закона за движение, следствия от тях и една схолия (поучение).
Първите, три определения са за маса, импулс (количество на движение) и инертност. Четвъртото определение е за сила. Следващите определения, от петото до осмото, се отнасят за центростремителна сила. Като примери на центростремителна сила Нютон привежда силата на тежестта, магнитната сила, онази сила (независимо от нейния произход!), която удържа планетите на криволинейните им орбити. От тези примери Нютон е показал, че е възможно да се „изстреля“ изкуствен спътник на Земята или да се отправи тяло от Земята, което да продължава вечно своето движение в небесното пространство. Тези две възможности са могли да станат действителност едва три века по-късно! Осмото определение е за ускорението.
След осемте определения следва знаменитото „поучение”, послужило като предмет на дълбоки размисли и дълги дискусии първоначално за философи, а след това и за физици, до наши дни. В това поучение са постулирани абсолютното време и абсолютното пространство, господствали във физиката до 19 век. За Нютон силата е абсолютен елемент (абсолютна величина), докато движението може да има само относителен характер поради отсъствието на абсолютна отправна система.
След определенията следват трите закона на движение: законът за инерцията, законът за пропорционалността на силата на ускорението и законът за действието и противодействието.
По-нататък Нютон разглежда движението на тела под действието на централни сили. С помощта на много просто доказателство Нютон установява валидността на следната теорема (и обратната на нея): движението на материална точка под-действието на централна сила е в една равнина и става така, че площта, описана от радиус-вектора, е пропорционална на изтеклото време. Изложени са и други твърдения, а именно, че силите са обратно пропорционални на квадрата на разстоянията, ако квадратите на периодите на обикаляне са пропорционални на кубовете на разстоянията. Формулира се и обща теорема за движение по крива линия около център. Това са нови теореми в механиката, достатъчни за обяснение на емперичните закони на Кеплер и разширяващи областта на приложимост на новата динамика за движението на планетите.
Най-голямото достижение в Нютоновите „Начала“ е построяването на „системата на света, изложена в последната трета книга на съчинението. Нютон отначало изброява установените от наблюдения закони за движенията на планетите Луната, спътниците на Юпитер и Сатурн. Като използва изложените по-напред резултати, Нютон дава динамична интерпретация на тези закони по същество в онзи вид, в който това сега се прави, а курсовете по физика, и стига до извода, че във всички случаи централната звезда действа на планетата или планетата на спътника със сила, обратно пропорционална на квадрата на разстоянието. Централен пункт в тази част от „Началата“ е доказателството, че силата, удържаща Луната на нейната орбита — това е онази сила, която заставя телата да падат на повърхността на Земята.
Преодолявайки всички трудности по пресмятането на силата на привличане между материална точка и тяло със сферична Форма. Нютон стига до извода, че
„Привличането съществува за всички тела изобщо и е пропорционално на масата на всяко от тях.”
Провъзгласявайки този закон, Нютон се въздържа от уточняване причината за привличането:
„Причината на тези свойства на силата на привличане досега не можах да изведа от явления; хипотези аз не измислям (hypotheses non figo).“
По-нататък Нютон излага знаменитата теорема за това, че две сфери, състоящи се от концентрични хомогенни слоеве, се привличат така като че ли техните маси са съсредоточени в центъра на всяка сфера. Нютон обръща внимание и на твърдението, че приливите и отливите на моретата стават от съвместното действие на Луната и Слънцето. „Началата“ завършват с формулирането на т. н. задача на Клеро, наречена по-късно „задачата за три тела“ и причинила толкова грижи и мъчнотии на математиците, започвайки от Нютон и стигайки до наши дни. Задачата се заключава в определяне движението на три тела (при Нютон — Слънце, Земя и Луна) под действието на централна (гравитационна) сила.
До Айнщайн гравитацията е била необяснимо явление в науката. Сам Нютон считал за безсмислено действието от разстояние без помощта на посредник (поле), но както бе споменато по-горе, винаги се отказвал публично да изрази своето отношение към природата на силата на тежестта.
Развитието на механиката и астрономията през 18 и 19 век са фактически триумф на Нютоновата механика. Не случайно Лагранж е казал, че Нютон е най-щастливият от всички световни гении, понеже системата на света може да се установи само един път.
Нютон е бил син на своето време, живял е с интересите на своя народ, на своята класа. Убеден привърженик на позицията „Законът е над краля“ — Нютон е бил избран в учредителния парламент. От януари 1688 г. до 1690 г. Нютон е в Лондон и взема участие в работата на Парламента. Тези години на интензивен живот, които заедно с наличието на несъмнена преумора, предизвикана от огромната работа по създаването на „Началата“, рязко влошили здравословното състояние на Нютон. Случаен удар — пожар, който унищожил редица книги на Нютон, го довели до нервно сътресение. Благодарение на грижите на неговите приятели Нютон бил излекуван и в 1694 г. пристъпил към работата върху теорията за движението на Луната. През 1696 г. Нютон бил назначен за пазител на Монетния двор. Кеймбриджският период от живота на Нютон завършил, започнал последният, лондонският период — период на обществено признание на заслугите на Нютон и славата му приживе.
В Лондон Нютон започнал да взема непосредствено участие в работата на Кралското дружество. На 30 ноември 1703 г. той бил избран за Председател на дружеството и останал такъв до края на живота си. В 1701 г. Нютон публикувал кратко съобщение: „Върху скалата на степените на топлината и студа“, което е една от основоположните работи в историята на термометрията. В 1704 г. излязла „Оптиката“ на Нютон. Тя била съставена от работите, написани още в Кеймбридж. „Оптиката“ донесла голяма слава на нейния автор. През живота на Нютон тя излязла три пъти: 1704 г., 1717 г. и в 1721 г. В 1706 г. бил издаден нейният латински превод, а в 1720 г. — френският. Към първото издание били приложени два математически трактата: „Върху квадратурата на кривите“ и „Класификация на кривите от трети ред“. Тази публикация дала повод за полемика с Лайбниц във връзка с приоритета. Полемиката продължила до смъртта на Нютон и създала както на Нютон, така и на Лайбниц не малко горчиви минути. (Сега е установено, че двамата автори дошли до откриването на диференциалното и интегралното смятане независимо един от друг, обаче Нютон направил откритието си няколко години по-рано от Лайбниц.)
„Оптиката“ се състои от три книги (подобно на „Началата“). В първата се разглеждат отражението, пречупването и дисперсията на светлината (анализ и синтез на цветове) с приложение за обяснение на дъгата и с допълнение, посветено на телескопите. Във втората книга се разглеждат оцветяванията на тънки слоеве. Накрая, третата книга, съдържа кратко експериментално изследване на дифракцията и завършва с 31 „въпроса“ от теоретически характер.
Книгата започва с определение на светлината, от което става ясно, че Нютон се е придържал към корпускулярната концепция за природата на светлината. След това се привеждат експериментите по дисперсията на светлината, които са станали основа на съвременната физическа оптика. Би било излишно да се подчертава гениалността на постановката на проблема, на изкусността на Нютоновото решение, на точността на измерването. Достатъчно е само да споменем грамадния скок, станал под влияние на работите на Нютон в изследванията на пречупването в призма, с които са се занимавали до него твърде много физици. Серията опити, които Нютон е направил, му позволили да заключи, че „Лъчи, отличаващи се по цвят, се различават и по степента на пречупване.“ и „Всяка хомогенна светлина има собствена окраска, отговаряща на степента на нейната (на светлината) пречупваемост, и такава окраска не може да се изменя при отражения и пречупвания.“ По-нататък Нютон привежда серии от опити, в същата стенен изумителни, относно синтеза на цветовете.
Всички тези открити от Нютон свойства на светлината му позволили да даде ново, по-пълно обяснение на дъгата и да изтълкува цвета на телата като резултат от избирателно поглъщане на падащата върху тях светлина.
В първата част на втората книга на „Оптиката“, състояща се от четири части, се описват серия от основни опити, проведени с изключително изкуство и станали класически. Тази част от работата е истински шедьовър на експерименталното изкуство. Нютон изучава оцветяването на тънки слоеве, като наблюдава това явление не само в бяла светлина, но и в монохроматична. Многобройните опити върху това интересно явление и точните измервания позволили на Нютон да открие различни закономерности, останали верни до наши дни.
Първата част от третата книга на „Оптиката“ съдържа няколко експериментални изследвания на явления, открити от Грималди. Обаче Нютон се старае да избегне думата „дифракция“. Както по подход, така и по интерпретация тези негови опити силно отстъпват на описаните в първите две книги и са подбрани с цел да представят явлението дифракция като резултат на привличане на светлинните корпускули от веществото: светлинните лъчи, преминаващи близо до краищата на телата, изпитват привличане и затова се изкривяват. Нютон чувствува недостатъчност в тази част от работата и в експериментално, и в теоретично отношение и заключава с честното признание, че не е успял да извърши онова, което е бил запланувал. Затова той предлага 31 „въпроса“ за по-нататъшно изследване, което следва да се направи от други учени. В действителност тези въпроси се отнасят не само до оптиката, но и до гравитацията, химията и други частни явления. В тях са събрани най-различни съображения на Нютон, в които наред с много дълбоки мисли се срещат и грешки, и очевидни противоречия. Известно е, че по-нататък, през 18 и 19 в., главно благодарение на опитите на Юнг и Френел във физиката се налага вълновата теория на светлината. Обаче сто години по-късно станало ясно, че е необходимо да се приеме и схващането на светлината като поток от частици. Интересно е, че в 1675 г. Нютон е разработвал теория на светлината, в която се съчетавала представата за вълните с представата за частици, запазващи своите „първоначални“ свойства. Твърде интересна е също така мисълта на Нютон за възможно превръщане на веществото в светлина и обратно. И наистина откритите през 1933—34 г. превръщания на електрон и позитрон в светлинни кванти потвърждават тази идея. По такъв начин Нютон предугадил бъдещо откритие във физиката, преминавайки в своите оптически открития пътя от шлифовката на стъкла и огледала (при конструирането на телескопите) до дълбоките теоретични разсъждения върху природата на светлината.
Краят на живота на Нютон е отбелязан със слава признание и спокойствие. В Кралското дружество той се ползвал с неоспорим авторитет, учениците му се сражавали за неговите идеи, за победата на Нютоновата физика по целия свят. Нютон е бил обкръжен с внимателните услуги на племенницата си. Нютон починал през нощта на 20 срещу 21 март 1727 г. Погребали го с големи почести в Уестминстерското абатство — английския национален пантеон — до гроба на друг велик син на Англия — композитора Хенри Пърсел.
Надписът на паметника над гроба на Нютон завършва с думите: „Нека смъртните се радват, че е съществувало такова украшение на човешкия род“.
На статуята на Нютон в Кеймбридж е изсечен следният стих от Лукреций: „С разума си той превъзхождаше човешкия род“.
И най-сетне в къщата, където той се е родил, е поместено двустишието на английския поет Александр Поп: „Природата и нейните закони били покрити с мрак. И рече бог: да бъде Нютон — и настана светлина“.
В „Мемоари“ („Memoirs“) на Академията на науките на Санкт Петербург от 1810 г. известният историк на науката Юлиус Клапрот (1783 – 1835) превежда „Признанията на мирния дракон“ на Мао Хуа – книга, донесена наскоро от Китай от френски пътешественик. В тази книга, датираща от 756 г. след Хр. (от времето на династията Тан), Хуа нарича кислорода „чи йин“ и обяснява, че макар той да не се среща никъде в чиста форма, се намира във въздуха. При горене се свързва с други елементи, включително метали, и образува нови вещества. Книгата добавя, че кислородът е една от съставките на водата. Ако Мао Хуа е открил кислорода през VIII век, това е постижение, което заслужава голяма слава. За съжаление няма следа от него, нито от неговата книга с изключение на работата на Клапрот. Ако Клапрот е сбъркал в датировката, може да се предположи, че Мирният дракон е направил тези признания, след като английският теолог Джоузеф Пристли (1733 – 1804) е открил кислорода. През 1774 г. Пристли публикувал по-пълно от това на Мао Хуа описание на кислорода. Ако всички участници са се движели бързо, откритието на Пристли може да е пренесено в Китай и обяснено на Мао Хуа, записано в един или няколко екземпляра, един от които случайно попаднал на французин, посетил Китай, който от своя страна го оставил в Русия навреме, за да успее Клапрот да подготви своята публикация от 1810 г. Такава верига от събития, макар все пак да е възможна, не може да обясни годината 756, цитирана от Клапрот. Мао Хуа, или който и да е този, който му е разказал за кислорода, остава една от най-загадъчните личности в историята на науката. Ако това се окаже фалшификат, тогава заслугата за първото откритие не е на Пристли, а на шведа Карл Шееле, който открил кислорода през 1772 или 1773 г. и публикувал своето откритие през 1779 г.
Михаил Василевич Ломоносов (1711 — 1765 г.). Роден е на 19.Х1.1711 г. в село Денисовка, Архангелска губерния (по-късно село Ломоносово, Архангелска област).
Епохата, в която се ражда и твори Михаил Василевич Ломоноеов, е епоха на бурно икономическо и културно развитие на феодално-крепостна Русия. Амбицията на Петър Велики да изведе Русия на едно от първите места между европейските държави довежда до развитието на манифактурата, корабостроенето, металургията и търговията, за които са необходими специалисти от всички области на науката и техниката. Създават се специални училища и необходимата за тях литература, а в 1724 г. в Петербург е основана Руската академия на науките, в която по-късно учи и работи Ломоносов.
Суровата природа на беломорския север, където живеят будни и предприемчиви хора, непознали татарското робство, помещичесхото земевладение, оказва извънредно благотворно влияние при формиране характера на Ломоносов, а културният и обществено-икономическият подем на Русия — върху творчеството му.
Още 10-годишен Михаил придружава баща си в пътуванията му из Бяло и Баренцово море, наблюдава многобройните и интересни явления на северната природа и натрупва много познания по морското дело и риболова. На 14 години пише без грешки и селяните го търсят за написването на необходимите им документи. Но освен граматиката на Смотрицки и аритметиката на Магнитски в родното място се намират само религиозни книги. В края на 1730 година Ломоносов избягва от къщи и се отправя пеш към Москва, за да изучава латински език, на който, по думите на учителя му Шубин, са написани най-хубавите книги.
Първоначално в Цифирната школа, а по-късно в Славяногръко-латинската академия Ломоносов изучава основно латински и гръцки език, чете в оригинал съчиненията на старите гръцки и римски мислители, сам написва твърде голям курс по риторика и прави първите си стихотворни опити. Неизяснени остават най-интересните за него въпроси — естественонаучните. В края на 1735 год. идва заповед на Сената в Петербург да се изпратят десетина от най-будните младежи на Спаските училища да продължат обучението си в Петербургската академия. Между избраниците е и безкрайно щастливият Ломоносов. Тук обучението се води по нова програма лекциите са на по-високо ниво, изучава се математика, риторика, история, география, латински и немски език и танци. Интересни лекции по физика чете академик Крафт, провеждайки опити с нови и съвършени за времето си апарати.
По същото време Академията организира голяма експедиция за изучаване на Сибир, за която е необходим химик, запознат и с минното дело. Тъй като такъв човек в Академията няма, ръководството решава да изпрати за подготовка в Германия трима студенти — Райзер, Виноградов и Ломоносов.
Марбургският университет, в който постъпват тримата младежи, е един от най-старите в Германия и първият основан без помощта на папата, поради което, се ползва с по-голяма свобода. Централна фигура е Хр. Волф, който въпреки идеалистическите си схващания е добър преподавател, а университетската библиотека разполага със съчиненията на Галилей, Торичели, Мариот, Хюйгенс, Декарт, Бойл и др. Под ръководството на Волф Ломоносов подготвя и изпраща в Петербург две самостоятелни работи „За превръщането на твърдите тела в течни“ и „Физическа дисертация за различията между смесените тела, състоящи се в сцепление на корпускулите“. В тях изяснява свойствата на телата от собствена материалистическа позиция.
– Вторият период от престоя в Германия във Фрайберг при Хенкел, който, по думите на Ломоносов, „презира всяка разумна философия“, е период на интензивна самостоятелна работа и наблюдение в рудниците. Тях по-късно оформя в съчинението „Върху земните слоеве“. Освен това работи върху руския стих и неговите особености. Поради изострени отношения с Хенкел Ломоносов напуска тайно града, отива в Маргбург, оженва се за дъщерята на скромен пивовар, но не остава дълго при немското семейство, а се отправя към Амстердам и търси кораб за Русия. По пътя се среща с видния холандски металург и химик Крамер и известно време работи в химическата му лаборатория. В Дюселдорф става жертва на измама и е вербуван за войник на пруския крал, но успяза да избяга, което прави престоя му в Германия невъзможен, и през юни 1741 г. се завръща в родината.
В Петербургската академия на науките Ломоносов постъпва първоначално като специалист, а от 1.1.1742 г. е назначен за асистент по физика. По това време в Академията господстващо влияние имат чужденците начело с Шумахер, които пречат на самостоятелното развитие на руската наука и култура. Още първите години Ломоносов повежда борба срещу чуждото влияние, за което е изключен от състава на Академията и дори престоява шест месеца в затвора. Разбирайки, че за да прокара своите идеи, трябва първо да се стабилизира, Ломоносов подава молба за присъждане на професорско звание и през юли 1745 г. е избран за професор по химия. Въпреки избора Шумахер изпраща научните му трудове за рецензия на Ойлер. Получава се извънредно ласкав отзив. Оттогава между двамата учени се поражда дружба, която продължава до смъртта на Ломоносов.
През първия период на своята дейност Ломоносов провежда предимно физически изследвания—до 1748 г., когато успява да издействува създаването на химическа лаборатория и поставя основите на руската химия. Като учен-енциклопедист до края на живота си работи в най-различни области на науката: развива корпускулярната теория, в която въвежда понятията „корпускула“ и „елемент“, извънредно много приближаващи се до съвременната молекула и атом; разглеждайки материя и движение в неразривна връзка, дефинира закона за запазване на веществото и движението; отхвърля теорията за топлорода и обяснява топлината чрез движението на корпускулите; разглеждайки химията в неразривна връзка с физиката и математиката, поставя основите на физикохимията като наука; провежда редица изследвания в областта на електричеството, метеорологията, геологията, минното дело, металургията, географията и историята; разработва конструкцията на оптични уреди, инструменти и редица въпроси из областта на теоретичната оптика; открива наличието на атмосфера на Венера. В областта на литературата и изкуството Ломоносов има заслуги за чистотата на руския език и звучността на стиха, разглежда редица стилистични и граматични въпроси за преобразуване на руския език, въвежда нови понятия и думи, научна терминология, сам пише стихове. Не случайно сто години по-късно Белински го нарича „Петър Велики на руската литература“.
За една от главните задачи на своята научна и просветителска дейност Ломоносов смята повишаването на културата на руския народ, от чиито недра ще израснат бъдещите нютоновци и платоновци. През целия си живот той се бори за създаване на демократически университет и успява в това си начинание. Като началник на академичната канцелария раздвижва редица въпроси във връзка с научната подготовка на студентите, увеличението броя на печатните книги и др., популяризира своите естественонаучни разбирания и се бори за изграждане на правилен естественонаучен мироглед. Тази широка дейност му спечелва много врагове, особено между духовенството, които по времето на Екатерина II взимат връх. Почива на 15.IV. 1765г. самотен, но непримирен, все още с много идеи за бъдещето на руската литература и Академията. Погребан е в лаврата Александър Невски.
Кавендиш е изследвал и обикновения въздух, за да определи състава на атмосферата. Карл Шееле (1742 – 1786), един от откривателите на кислорода и азота, вече е определил, че въздухът е смес от двата елемента. Кавендиш е установил, че пропорцията е 20,83 % кислород и 79,17 % азот, като всичко друго се смята за примеси.
Водородът е открит през 1766 г. от състоятелния класически учен Хенри Кавендиш (1731 – 1810) в неговата частна лаборатория в Лондон. Той го кръстил „избухлив въздух“ заради това, което се случва, когато към него се поднесе огън. През 1781 г. Кавендиш открил, че може изкуствено да създаде вода, когато взриви заедно водород и обикновен въздух. По-късно повторил експеримента с водород и кислород и успял да изчисли, че водата е съставена от тези два елемента и да определи пропорциите на всеки от тях в нейния състав.
Шведският химик Йонс Якоб Берцелиус (1779 – 1848) е съставил първата таблица на елементите, в която всеки елемент е подреден в съответствие със силата на неговия положителен или отрицателен електрическа заряд. Той е открил, че големината на афинитета между два елемента е пропорционална на разстоянието на което са разделени те в неговата таблица.
Oткрити са от К. де Дюв в началото на 50-те год. На 20 век.
Лизозомите са органиоди и на вътрешната мембранна система на клетката, изпълняваща литични функции те участват в смилането на екзогенните вещества и в елиминирането на отделни вътреклетъчни части (автофагия). Имат кълбовидна, яйцевидна форма и големина от 0,1 до 0,8 µm. Изградени са от гранулирана елекртонноплътна маса, наречена матрикс обграден от двуслойна мембрана, която за разлика от митохондриалната не образува кристи.
Гранулираната вътрешност на йизозомите съдържа голям брой хидролитични ензими, между които значително място заема киселата – фосфатаза, която се смята като маркер на лизозомите. Лизозомите съдържат и други ензими – нуклеази, фосфопротеинфосфатаза и др. Количеството на хидролазите в лизозомите е огромно.
Лизозомите извършват и м-са на апоптолиза – самоунищожение на клетките.
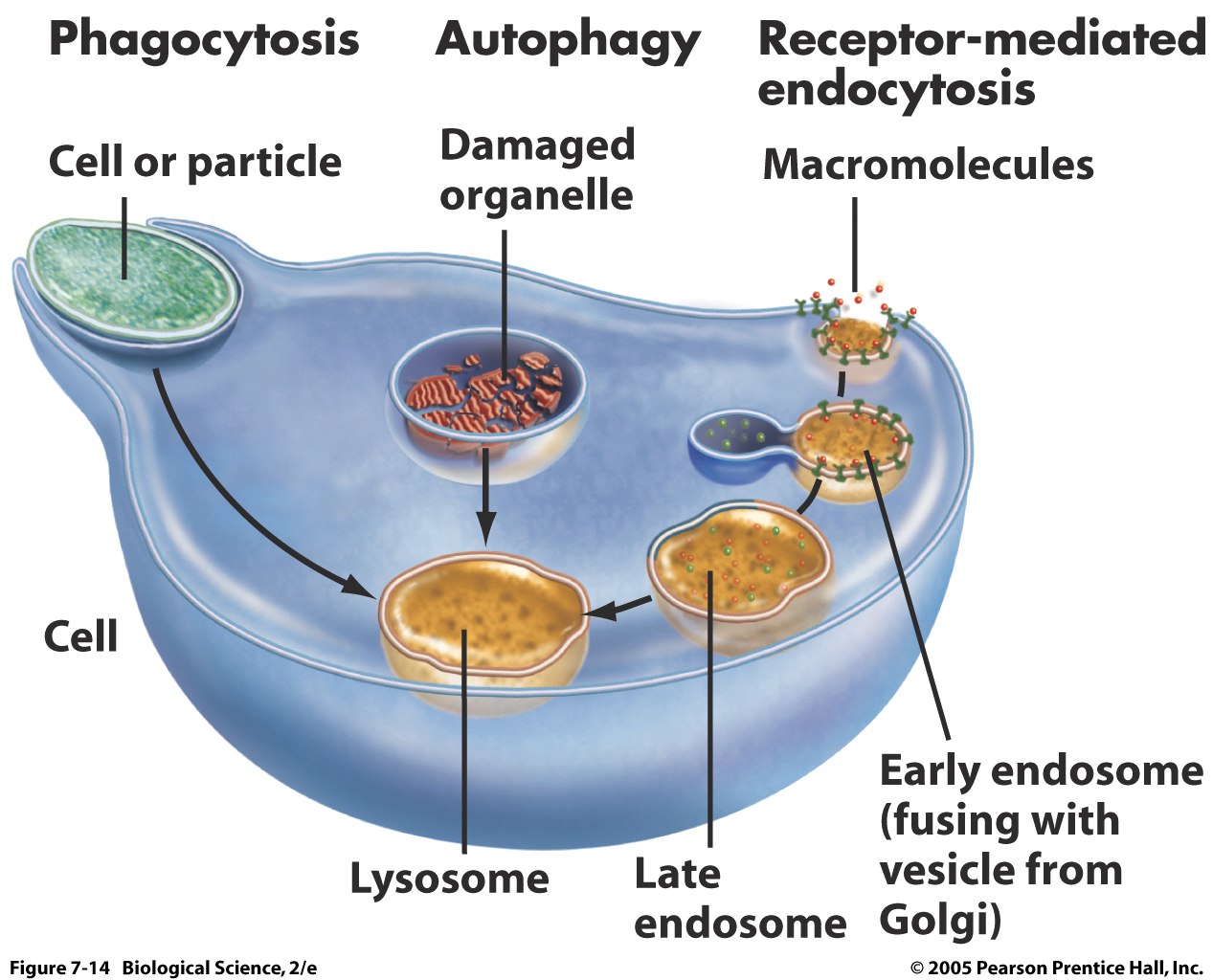
Лизозомите се подразделят на първични и вторични.
Първични лизозоми – Те са изградени от финно гранулиран плътен матрикс. Образуват се от периферните издути части на цистерните на апарата на Голджи. Първичните лозозоми не са активни, защото се съдържат събстрат ( чужд белтък, бактерии и др.)
Вторичните лизозоми са ативни, затова в тях често се срещат полъсмлени или неразтворени остатъци от чужди вещества или собствени вещества от метаболизма на клтката. Вторичните лизозоми се подразделят на :
Смилателни вакуоли
Автолизозоми
Щитоергозоми
Мултивезикуларни телца
Телозомни ( остатъчни телца)
Лизозома
ФУНКЦИЯ НА ЛИЗОЗОМИТЕ
Когато „остареят”, част от част от органелите на клетката (митохондрии, едноплазмен ретикулум и др.) се елиминират, като първоначално се ограждат от каналчета на едноплазмения ретикулум, изпълнени смного хидролитични ензими. Тези каналчета, съсдържат хурдрол. зими и огадените цитоплазмени органели образуват автофагозома. Това явление се нарича автофагия. Към тези тени автоагозоми се присъединяват първичните лизозоми и автофагозомите се превръщат в автолизозоми (цитолизозоми). Когато автолизозомите се слеят образуват цитосегрозоми. Когато съдържанието в автолизозомите, респективно в цитосегрезомите се разгради под действието на хидролит. ензими (и след като част пт смлените вещества се използват от клетката ) остават крайни образувания – телолизоми. Веществата от, които се изхвърлят от клетката след преработка.
Лизозомите (автолизозомите и цитосегрезомите) извършват и автолиз (според някой автори понятието е апоптоза) на клетката, ако тя е увредена.
Лизозомите имат и друга функция – модигикация на клетъчни продукти. Наример в клетките на штитпжидната жлеза едноплазмения ретикулум синтезира тироглобулин (белтък предшественик на тиропния хормон) След секрецията на тироглубина от едноплазмения ретикулум, последния се слива с първичните лизозоми, хидлолитичните ензими на които го трансформират в тироиден хормон, който след това се извежда в кръвния поток.
Според съвременните представи лизозомния апарат се е развил като специализирана част на катаболитната система на цитоплазмата с широка функционална пластичност.
ПЕРОКСИЗОМИ
Рази група микро телца е наречена пероксизоми от К. Де Дзв през 1956 г. Те са неголеми вакуоли, обвити с единична мембрана, ограничаваща рганулирания матрикс. В центъра на матрикса има сърцевина. В зоната на сърцевината се намира паракристална структура, състояща се от подредено опаковани фибрили и тръбици.
Пероксизомите са открити при протисти (амеби), при низити гъби (дрожди), при висши растения (ембрионални тъкани, ендосперм), при висши гръбначни животни, главно в черния дроб и бъбреците.
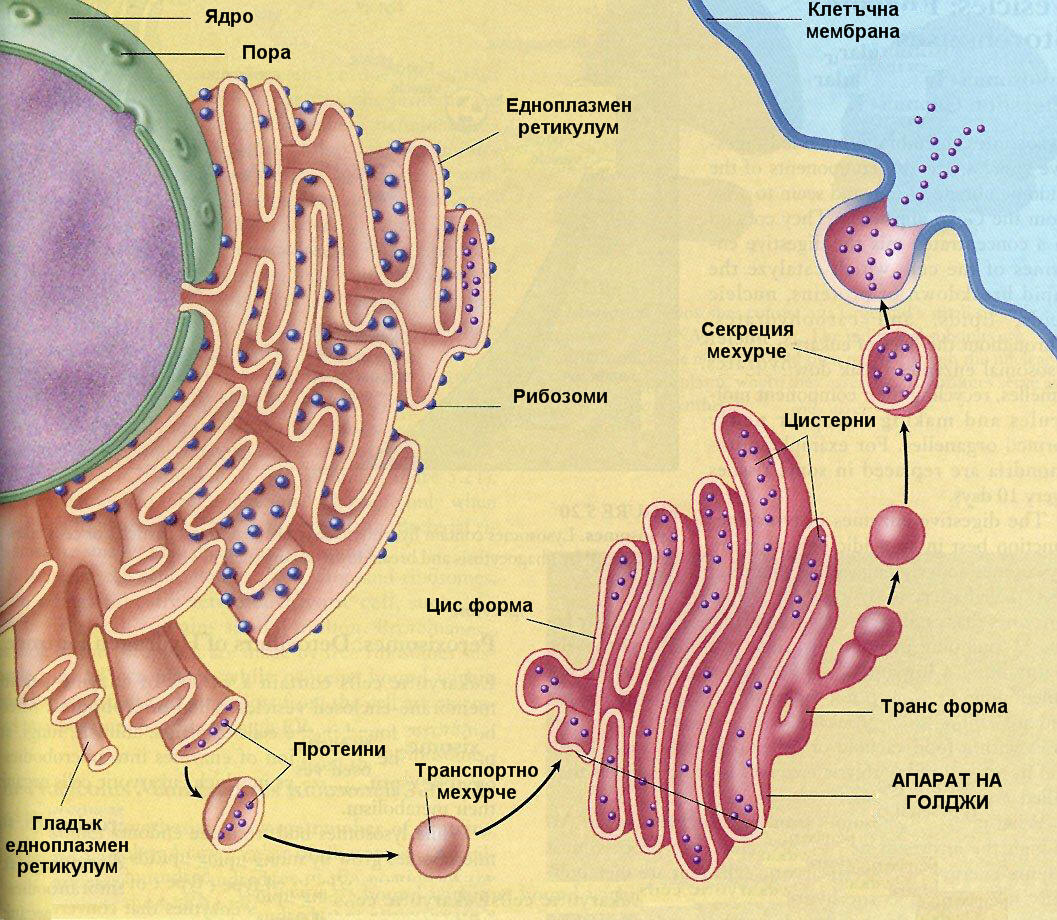
Колмплекса на Голджи е описан през 1898 г. от Камило Голджи. Той го е наблюдавал като мрежест венец около ядрото на нервни клетки.
Обикновено той се намира до ядрото на клетката или около него. Това е свързано с основната му функция – секреторна. Възможно е и промяна на локализацията му в някой клетки във връзка с физиологичните особености на клетката.
Комплекса на Голджи съществува в три нива на организация:
ЦИСТЕРНА – Сплесната мембранна торбичка с централно плочковидна част и перфорирана периферна зона.
ДИКЦИОЗОМА – Група от цистери (4-8), наредени в пакет.
Комплекс на Голджи – Няколко свързани диктиозоми. Броят им варира във връзка с физиологичните потребности на клетката.
Основна структурна единица на комплекса.
Дикциозомата е съставена от плътно разположени една до друга плоски мембранни цистерни, между които има тънък слой цитоплазма. Освен цистерните в зоната на дикциозомата се наблюдават множество вакуоли
АПАРАТ НА ГОЛДЖИ
Комплека на Голджи е полрна структура. Прието е да се различават проксимален (близък, цис) и дистален (далечен – транс) ползси. В секретиращите клетки проксималния край на комплекса е обърнат към ядрото (или едноплазмения ретикумум), а дисталния край към клетъчната мембрана (плазмената). Към проксималния полюс към цистерните на комплекса се доближават малки вакуоли (транзитни) – (транзитните вакуоли идват от едноплазмения ретикулум), а в дисталния край характерзиращи се с по-едрите разширения на цистерните, се образуват големите бакуоли съдържащи продукта на клетъчната секреция ( образувани от комплекса на Голджи)
В клетките отделните дикциозоми магат да бъдат свързани чрез система от цистерни и вакуоли.
Така се формира тримерна мрежа.
! Някой автори вместо понятието дикЦиозома използват понятието дикТиозома.
ФУНКЦИИ НА АПАРАТА НА ГОЛДЖИ !
Апарата на Голджи участва в натрупването и секресията на продуктите синтезирани в едноплазмения ретикулум, участва в тяхното химическо преустройство и узряване, полимеризация на полизахариди и образуване на комплекси от полизахариди (ПЗ) и белтъци (Б) – гликопротеиди; атно на комплекса му липиди(Л) и Б -липопротеиди и др. Освен тези вещества апарата на Голджи натрупва и други вещества – ензими, хормони, vit. Прието е, че в апарата на Голджи се извършва адсорбиране, кондензиране и опаковане на веществата, които се синтезират в гранулирания еноплазмен ретикулум и постъпват чрез транспортни вакуоли в апарата на Голджи. В апарата на голджи се формират и опаковат лизозомите и преоксизомите.
Комплекса на Голджи присъства във всички клетки на еукатиотните организми ( с изключение на еритроцитите при млекопитаещите организми), но не всяка клетка притежава способност за секреция на белтъци, Л и ПЗ (мускулни клетки, еритроцити, покривен епител). Всички те обаче притежават добре развити комплекс на Голджи. Това е свързано с образуването на лизозомите.
НЯКОЙ СПЕЦИФИЧНИ ФОРМИ НА АПАРАТА НА ГОЛДЖИ
ПРИ РАСТИТЕЛНА КЛЕТКА междуклетъчната пластинка, разделяща двете нови, дъщерни клетки след приключване на кариокинезата (делене на ядрото), се обр. от апарата на Голджи. Междуклетъчния плас е продукт на вакуоли от комплекса на Голджи, които изливат съдържанието си в екваториалната плоскост (пектин и хемуцекулата) на делящата се клетка.
Междуклетъчната пластина е изградена от пектин и хемицелулоза.
РАЗВИТИЕ НА АКРОЗОМАТА ПРИ ПРЕРМАТОЗОИДИРЕ
АКРОЗОМАТА Е МЕМБРАННА СТРУКТУРА на сперматозоида, с която той разпознава и свързва сперматозоида към повърхността на яйцеклетката.
Аузиращите от апарата на Голджи вакуоли увеличават повърхността на акрозомата.
Този сайт използва ‘бисквитки’ (cookies), за да ви предостави възможно най-добро потребителско изживяване. Можете да промените настройките си за бисквитки, или в противен случай приемаме, че сте съгласни с нашите условия за ползване.ПриемамПрочети повече
Правила на поверителност
Privacy Overview
This website uses cookies to improve your experience while you navigate through the website. Out of these cookies, the cookies that are categorized as necessary are stored on your browser as they are essential for the working of basic functionalities of the website. We also use third-party cookies that help us analyze and understand how you use this website. These cookies will be stored in your browser only with your consent. You also have the option to opt-out of these cookies. But opting out of some of these cookies may have an effect on your browsing experience.
Necessary cookies are absolutely essential for the website to function properly. This category only includes cookies that ensures basic functionalities and security features of the website. These cookies do not store any personal information.
Any cookies that may not be particularly necessary for the website to function and is used specifically to collect user personal data via analytics, ads, other embedded contents are termed as non-necessary cookies. It is mandatory to procure user consent prior to running these cookies on your website.

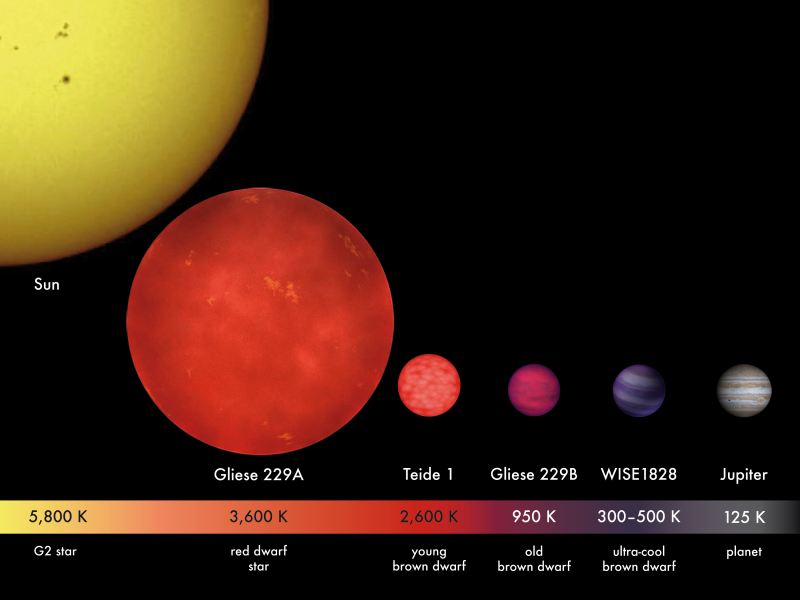
 Кафявите джуджета са малки звезди, чиято маса не е достатъчна, за да започне ядрено горене на водород в ядрото им. В резултат на това повърхностната им температура е само 1-2 хиляди градуса, а когато остареят температурата им пада до тази на планетите. За това те са тъмни и много трудни за откриване. Първата такава звезда е била открита преди 10 години.
Кафявите джуджета са малки звезди, чиято маса не е достатъчна, за да започне ядрено горене на водород в ядрото им. В резултат на това повърхностната им температура е само 1-2 хиляди градуса, а когато остареят температурата им пада до тази на планетите. За това те са тъмни и много трудни за откриване. Първата такава звезда е била открита преди 10 години. По-масивното от двете новооткрити джуджета се нарича HD 3651 B и се намира в съзвездието Риби. Обекта се намира в слънчева система, чието слънце е по-малко от нашето и бива обикаляно от доста по-малка от Сатурн планета. Орбитата на планетата е силно елиптична, което караше учените да мислят, че някой друг по-масивен обект придърпва планетата с гравитацията си навън. Това със сигурност е Т-джудже. Това е доказателство, че планетарни слънчеви системи могат да приютяват кафяви джуджета, които да правят орбитите на планетите доста странни.
По-масивното от двете новооткрити джуджета се нарича HD 3651 B и се намира в съзвездието Риби. Обекта се намира в слънчева система, чието слънце е по-малко от нашето и бива обикаляно от доста по-малка от Сатурн планета. Орбитата на планетата е силно елиптична, което караше учените да мислят, че някой друг по-масивен обект придърпва планетата с гравитацията си навън. Това със сигурност е Т-джудже. Това е доказателство, че планетарни слънчеви системи могат да приютяват кафяви джуджета, които да правят орбитите на планетите доста странни.
 Изображение:
Изображение:  Наполеон бил млад и идеалистичен, поради което се върнал в Корсика да брани независимостта на своята малка родина. Скоро обаче той обърнал гръб на родното си място и се отдал на Франция. Това станало след като участвал в неуспешна атака в Сардиния.
Наполеон бил млад и идеалистичен, поради което се върнал в Корсика да брани независимостта на своята малка родина. Скоро обаче той обърнал гръб на родното си място и се отдал на Франция. Това станало след като участвал в неуспешна атака в Сардиния.



 Михаил Василевич Ломоносов (1711 — 1765 г.). Роден е на 19.Х1.1711 г. в село Денисовка, Архангелска губерния (по-късно село Ломоносово, Архангелска област).
Михаил Василевич Ломоносов (1711 — 1765 г.). Роден е на 19.Х1.1711 г. в село Денисовка, Архангелска губерния (по-късно село Ломоносово, Архангелска област).
 Кавендиш е изследвал и обикновения въздух, за да определи състава на атмосферата. Карл Шееле (1742 – 1786), един от откривателите на кислорода и азота, вече е определил, че въздухът е смес от двата елемента. Кавендиш е установил, че пропорцията е 20,83 % кислород и 79,17 % азот, като всичко друго се смята за примеси.
Кавендиш е изследвал и обикновения въздух, за да определи състава на атмосферата. Карл Шееле (1742 – 1786), един от откривателите на кислорода и азота, вече е определил, че въздухът е смес от двата елемента. Кавендиш е установил, че пропорцията е 20,83 % кислород и 79,17 % азот, като всичко друго се смята за примеси.")