Още преди появата на цивилизацията първобитните пастири са забелязали, че животните наследяват характерни белези от родителите си и че след няколко поколения желаните характеристики могат да бъдат затвърдени.
Изкуствена селекция
Ползвайки наученото, те са видоизменяли своите стада, като са отстранявали неподходящите животни и са получавали потомство само от избрани индивиди. Този еволюционен процес на изкуствена селекция ни е дал овцата (получена от дивата коза), кравата (произлязла от бизона) и прасето (произлязло от глигана).
Естествен подбор
Идеята, че подобен процес на естествена селекция е довел до развитието на добре приспособени видове, обитаващи днешния свят, е застъпена за пръв път в съчинението „Върху природата“ (V в. пр. Хр.) на гръцкия философ Емпедокъл от Акрага, автор на теорията за големия взрив. Неговата представа за еволюцията е твърде незряла – той предполага, че сред изчезналите, зле приспособими видове, изхвърлени при естествения подбор, може да е имало такива недоразумения като същества с глави, но без вратове или пък с ръце, но без рамене. По-късно гръцките мислители съставят значително по-добре обмислена теория. Естественият подбор потвърждава тяхната теза, че добре приспособените организми съществуват, защото сред случайно създадените при естествения градивен процес продукти те са успели най-добре да оцелеят и да се размножават. За поддръжниците на тази теория тя отрича вярването, че отделните същества са измислени и лично изработени от Бог. През I в. пр. Хр. естественият подбор отново намира своето място в „За природата на нещата“ от Лукреций:
„Преди един вид да оживее – той се нуждае от храна преди всичко и от части за възпроизводство,
чрез които семената на живота могат да намерят своя път
от мъжа към жената и да съединят телата
във взаимно удоволствие.
През онези стари дни
много опити бяха неуспешни, много от видовете
не успяха да оцелеят; това, което виждаме сега
да се радва на дъха на живота, трябва от първите
да е намерило защита за своя характер,
за своята сръчност, смелост и бързина.“
Идеите в това стихотворение са на гръцкия философ Епикур. Неговите научни схващания са заимствани най-вече директно от Демокрит и Левкип, създателите на теориите за атомите и молекулите. Вероятно един от тези двама гърци, живели през V в. пр. Хр., е първият, който е дал убедително обяснение за естествения подбор. Но тъй като техните трудове са изгубени, не можем да сме сигурни.
Произход на човека от маймуната
Големият ислямски учен Абу Рейхан Мухамед Ибн Ахмед ал-Бируни от Хорезм (973 – 1048) стига до заключението, че човекът е родствено свързан с други по-примитивни биологични видове. В своята книга „Китаб ал-джамахир“ той пише, че човечеството е резултат от еволюцията на дълга верига бозайници, най-близки от които са приматите (човекоподобните маймуни).
Еволюция на съвременния човек
Възгледът, че човешкият род е еволюирал от първобитния човек до днешния Homo sapiens, е бил изказан най-напред в Китай в сборника с научни и философски трактати „Хуай Нан Дзъ“ (II в. пр. Хр.), издаден в Нан Дзъ от Лю Ан по времето на династията Хан. Според тази книга човекът е минал през пет етапа в своята еволюция – започвайки от загадъчния „димен човек“ и достигайки до съвременния „обикновен човек“.
Дегенерация
Знанията за същността на еволюционния процес водят до появата на специфични възгледи относно бъдещето на човешката раса. Един от тях е политиката на евгеника, застъпвана от Платон. Според друг възглед многократното размножаване вътре в ограничен род води до дегенерация и непригодност. Тази идея се появява през 1813 г. в Англия в книгата „Лекции по физиология, зоология и естествена история на човека“, в която се твърди, че европейските управляващи среди дегенерират и е необходима тяхната подмяна. Английският председател на Камарата на лордовете не одобрява книгата я и обявява за противоречаща на Светото писание. Нейният автор – хирургът Уилям Лоуренс – се отказва от нея, за да запази престижа си. Но по-късно той е награден със званието барон и получава поста на старши хирург на кралицата, а Чарлс Дарвин преповтаря много от неговите изводи за естествения подбор в своето съчинение „Произход на видовете“ (1859). Дарвин не отдава нужното на Лоурънс, но признава, че еволюционната теория, развита в „Произход на видовете“, е много стара: „Много далеч от истината би било да се твърди, че този принцип е открит в днешно време… Принципа на селекцията аз видях ясно записан в една древна китайска енциклопедия.“
Останки, открити в Зави Чеми Шанидар в Северен Ирак, разкриват, че животновъдството е било практикувано от номадите по тези земи от около 9000 г. пр. Хр. Те вече отглеждали предшественицата на днешната овца,макар че тя все още била доста близка родственица на козата. Вероятно нейното късо право руно не е било подстригвано и тя трябва да е била развъждана за месо и мляко, а не за вълна. (Макар че облеклото може да е било правено от кожата на овце.) Така че, освен ако в бъдеще не се появят по-стари открития, изглежда, че отглеждането на добитък е започнало в тревните площи на Западна Азия през X хилядолетие пр. Хр. Подходящите терени са били разположени в Иран, Ирак, Южна Турция, Сирия и Палестина.
Ветеринарни пестициди
Те са известни от книга отпреди около 3000 години. Един от тях е служел за изкореняването на вътрешни паразити като чревни глисти, като се използват противоглистни растения.
Дезинфекционен разтвор за унищожаване на вредители в руното на овцете се е използвал още в древен Картаген (днешен Тунис). Руното се обработвало със специално масло, открито в извор в околностите на града, което имало забележителен мирис на стърготини от лимоново дърво. Практическото му използване започнало може би около средата на I хилядолетие пр. Хр.
Земеделието, най-важната технологична революция в света, започнала значително по-късно, вероятно около 8000 г. пр. Хр., някъде между средиземноморския бряг и Южен Иран. Следи от култивирано жито и ечемик, датиращи към 7500 – 6500 пр. Хр., са открити в днешна Турция, Палестина и Иран. Тяхното широко разпространение показва, че култивирането на посевите е започнало значително по-рано.
Наръчник по земеделие
Около XVIII в. пр. Хр. в Месопотамия било съчинено стихотворение на шумерски с указания как да се гледа ферма. Стихотворението е достигнало до наши дни, написано с клиновидно писмо върху осем глинени плочки от град Нипур. То започва с инструкции за напояване и стига до инструментите, почвата, оранта и сеитбата, грижите за посевите, жътвата и вършитбата.
Анализ на почвата
Най-старата китайска книга в проза „Шу Джин“ или „Документална класика“ включва описание на типични почви от деветте провинции, на които бил разделен Северен Китай. Първият документ в раздела на книгата, отнасящ се до династията Хсия (около 2100 -1700 г. пр. Хр.) описва стопанските особености на Китай, провинция по провинция, включително състава на почвата и често цвета и хранителността. Това е много кратък стар документ, съдържанието на който може да се е предавало устно около 2000 г. пр. Хр. и след това да е било записано
Земеделски пестициди
Опушване за унищожаване на насекоми е използвано за пръв път в Северен Китай в началото на династията Джо преди 1000 г. пр. Хр. Спрейове, аерозоли, химикали за ветеринарството и други продукти се появяват по-късно. Официалните отговорници по дезинфекцията във времето на династията Джо имали между другите си задължения да контролират жабите, водните вредители и насекомите.
Веялка
Веялката за механично отделяне на зърната от плявата е изобретена в Китай около III в. пр. Хр. (най-ранните останки, включително макети, са от първия период на династията Хан). Веялката се състои от въртяща се в калъф перка, която тегли въздух през отвори около оста си и го издухва по хоризонтална тръба. Зърната се подават във вертикална шахта, пресичаща тръбата, и когато попаднат във въздушната струя, люспите се отвяват. Зърната падат през отвор до веялката, а люспите се отвяват в края на тръбата.
Подвижни плугове
Робърт Бойър Смит имал малка ферма на полуостров Йорк, Южна Австралия. Земята била камениста и орането било кошмар. Заедно с брат си Клерънс (ковач) той изобретил подвижния плуг и демонстрирал първия модел през 1876 г. Три рала, свързани едно с друго, дълбаят паралелни бразди. Всяко е конструирано така, че да се вдига и обхожда препятствието, което срещне, след което от тласъка на тежестта заема отново позиция за оран. Когато парламентът на Южна Австралия решил да награди изобретателя на плуга с пари и земя, възникнал спор кой от братята заслужава да бъде възнаграден. Накрая през 1882 г. бил предпочетен Смит.
Плуг
Преди да бъде създаден тракторът, най-големият прогрес в увеличаването на ефективността на земеделския инструментариум бил постигнат с плуга. Чрез ограничаване на триенето значително се намалява силата, необходима за теглене на ралото, така че едно животно може да върши работата на дузина. Писателите и художниците не намират вдъхновение в земеделието, така че живописта и литературата не откриват нищо в революцията на орането. Но в Китай са намерени много железни плугове от периода на династията Хан (206 г. пр. Хр. – 9 г. след Хр.). Тъй като тяхната форма е твърде усложнена, ясно е, че плоските дървени плугове (които не са оцелели през вековете) са от по-раншния Период на непримиримите държави (481 – 221 г. пр. Хр.). Те вероятно са използвани през 300 г. пр. Хр.
Редосеялка
Редосеялката е създадена от шумерските земеделци през III хилядолетие пр. Хр. в Южна Месопотамия. Ралото се състои от един, а може би понякога от два или три зъбеца и резервоар за семената на височината на гърдите, откъдето през тръбички семената се отвеждат към отворите, които се нагласяват така, че семената да могат да бъдат засаждани на оптимална дълбочина в почвата. Редосеялките се теглят от волове. Работата се извършва от четири човека. Един от тях има задачата да поддържа необходимото количество семена по време на работата с редосеялката. Според първия учебник по земеделие (по-горе) за земеделеца било важно да контролира този процес, както и да коригира дълбочината, на която се оре.
Наука за почвата
Научният анализ на почвата започва да се прави през IV в. пр. Хр., когато се появява в глави от „Гуан Дзъ“ – книга, съставена главно в източнокитайската провинция Чи. Една глава от „Гуан Дзъ“ представлява анализ на почвата област по област, основан на дълбочината на горната и долната граница на подпочвените води, топографията, състава, цвета, ботаниката, преминаването на звука през суха почва, ефекта върху човешкото здраве и солеността. Плодородните почви са класифицирани с помощта на обективна скала от едно до десет.
Пестициди за семената
Най-ранното регистрирано третиране на семената с пестициди било в Гърция през V в. пр. Хр. Говори се, че философът Демокрит е препоръчвал семената от зеле да се накисват в сок от дебелец (ушно биле). Пестициди за обработка на житни семена трябва да са се появили не много по-късно и на Запад, и в Китай. Те се споменават в китайска книга по земеделие от I в. пр. Хр. (където като токсично вещество е посочен аконит – самакитка) и в „Естествената история“ на Плиний, публикувана в Рим през I век. И в двете книги вероятно става дума за препарати, които вече се се използвали много преди появяването на самите книги.
Биологичен контрол на вредители
Биологичният контрол на вредители нормално означава използването на един вид, за да се намали друг, Обаче най-ранният регистриран пример за биологичен контрол е използването на видовете за собственото им ограничаване. В Юдея (днешен Израел) по римско време, около 100 г., хората започнали да събират мравки от мравуняците им и да ги слагат в други мравуняци, При това различните групи мравки, разпознавайки се като чужди, започвали да се избиват помежду си. Това е доста трудоемък и не особено практичен начин за ликвидиране на мравките. Биологичният контрол с използване на един вид срещу друг бил въведен през III в., за да се запазят мандариновите насаждения в Южек Китай. На пазарите в Джао Джи се продавали гнезда на цитрусови мравки в торби от рогозка. Земеделците ги прикрепяли към дърветата, за да държат настрани другите насекоми, които са опасни за реколтата, не представляват храна за лимоновите мравки.
Книгата на Сю Йоу „Описание на някои традиции на математическата практика“ от 190 г. след Хр. описва системата на „великото единство“ за графично представяне на числата, преподавана на автора от Тиен Му Сиен Сен, който е описал и сметалото. Великото единство е графика, на която оста X (хоризонталата) е градуирана в степени на десет, т. е. единици, десетици, стотици, хиляди, милиони, милиарди и т.н. Оста V, или вертикалната ос, е градуирана в единици от едно до девет. Така в този предвестник на Картезианската аритметика всяко число може да се представи графично, като се движат броячите по деветте хоризонтални линии, започващи от оста Y.
Нов метод за анализиране на геноми подтвърждава, че неандерталците са се кръстосвали с предците на евроазиатците.
Техническите възражения срещу идеята, че неандерталците са се кръстосвали с предците на евроазиатците бяха преодоляни, благодарение на нов метод за анализиране на генома. Техниката може по-сигурно да засече генетичните следи на кръстосването, отколкото предишните методи за анализ и ще бъде използвана за изследвания на други стари или редки ДНК проби.
“Нашият метод може да разграничи двa случая с едва доловими разлики, които могат да обяснат генетичните прилики, споделени между неандерталците и модерните хора от Европа и Азия” – каза съавторът на изследването Конрад Лохзе, популационен генетик в Единбургския университет. Първата възможност е, че неандерталците редовно са се кръстосвали със съвременните хора, след като емигрирали от Африка. Алтернативният варианте, че хората, които напуснали Африка еволюирали от същата потомствена субпопулация, от която произхождат неандерталците.
Много изследователи спорят, дали кръстосването между видовете е по-вероятно. Това е така, защото отговаря на генетичния модел, наблюдаван в проучвания, които сравняват геномите на много съвременни хора. Но новият подход напълно изключва алтернативния вариант, без да изисква допълнителна информация, използвайки само тази, извлечена от един геном от всеки вид: неандерталец, европеец/азиатец, африканец и шимпанзе.
Същият метод може да бъде полезен в други проучвания за кръстосване, където малко проби са на разположение. “Тъй като методът използва максимално информацията в индивидуалните геноми, особено вълнуващо е използването му за разкриване на историята на видове, които са редки или изчезнали.” – каза Лохзе. Всъщност, авторите първоначално развивали метода, изучавайки историята на популации на насекоми в Европа и островни видове прасета в югоизточна Азия, някои от които са изключително редки.
Лохзе е против да се отделя прекалено внимание на факта, че новият метод предполага малко по-високгенетичен принос на неандерталците за съвременните хора, отколкото минали проучвания. Оценяването на този принос е сложно и е възможно да варира взависимост от подхода.
“Тези резултати са важни, защото запълват пропуск в спора за това, дали неандерталците са се кръстосвали с хората. Методът може да бъде използван за разбирането на еволюционната история на други организми, включително и видовете, застрашени от изчечване” – каза Марк Джонстън, главен редактор на списанието Genetics.
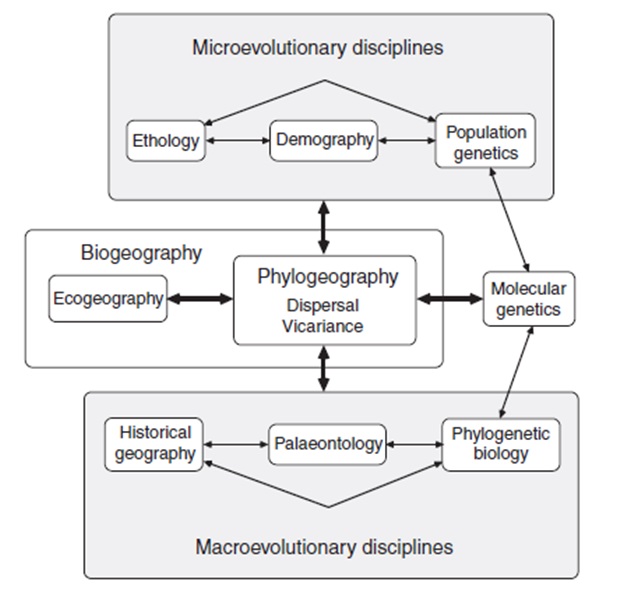
Филогеографията е нова дисциплина, обединяваща няколко дисциплини като генетика, молекулярна билогия, еволюционна биология, зоология, биогеграфия, ботаника и др. Стартираме с една преводна стария на човека дал името на тази дисциплина – Джон Авайз.
Джон Чарлз Авайз, роден през 1948 година е амеркански еволюционен генетик, еколог, консервационист и природен историк. Професор по Екология и еволюционна биология в Калифорнийския университет в Ървин. Той пръв дава името на новата дисциплина филогеография през 1987 година, като първите изследвания които прави в неговата лаборатория са посветени на генетичното родство и размножителното поведение при рибите и други животни.
Д-р. Чавдар Черников
/John C. Avise*.Phylogeography: retrospect and prospect. Journal of Biogeography (J. Biogeogr.) (2009) 36, 3–15/
Превел: Димитра Лефтерова
Въведение
Филогеографията е относително нова дисциплина, която се занимава с пространствения аранжимент на генетичните линии, по – специално между близкородствени видове. Името филогеография е дадено през 1987 (Avise et al., 1987). Приблизително десетилетие по – рано учените използват митохондриална ДНК за да изяснят какблизкородствени видове са генеалогично свързани с вероятен общ предшественик/ митохондриалния геном е много подходящ за тази цел/. При генетични изследвания на различни видове са разкрити точни начини на пространствена подредба на мт ДНК генетични линии. Така да се каже, генеалогия се свързва с географията. След 1987 г., филогеографията става удобен способ за решаване на генеалогични въпроси на пространствените и времевите параметри намикроеволюцията. Във физиката Теорията на относителността на Алберт Анщайн ни дава концепцията, че времето и пространството са взаимосвързани и изкривяванията на времево – пространствения континуум могат предсказуемо да повлияват пътя на свободните частици. При филогеографията, няма подходящи математични закони, определящи точно начина на пространствената конфигурация на генетичните линии/ еволюционните процеси са твърде вариабилни и индивидуално различни за това/. Въпреки това за немалко видове сравнително филогеографски изследвания разкриват демографската и историческата природа на вътревидовата еволюция. Започвам този преглед с някой вече стандартни описания за изследвания на цитоплазмения геном и генногенеалогичната теория (Hudson, 1990), които най – често се използват при филогенетичните изследвания. След това ще илюстрирам, чрез някои практически примери ключови открития и новопоявили се принципи на филогеографията, преди да завърша с някои мисли за бъдещите насоки на тази процъфтяваща дисциплина. Тази статия предлага въвеждащи основи в областа, за повече и по – подробни обяснения и по– обширни цитати може да разгледате първата филогеографска литература Avise (2000).
Молекули и анализи
Митохондриалната ДНК еволюира бързо при животните
Митихондриалната ДНК ни връща обратно към протобактерия, която се е сляла ендосимбиотично с пре – еукариотна клетка рано в историята на живота на земята. По време на еволюцията, този цитоплазмен геном е загубил голяма част от своятаДНК и сега типично е представен в животинската клетка само с 37 гени всички свързани в една циклична молекула с приблизителна дължина от 17 000 нуклеотидни двойки. Два от тези гени кодират рибозомни РНК-и, 22 специфично различни транспортни РНК молекули и 13 кодират белтъчни субеденици които се колаборират с нуклеарно кодирани белтъци в биохимичните пътища на електронния транспорт и окислителното фосфорилиране, което помага за генериране на клетъчната енергия. Молекулата на мтДНК също така включва и контролен регион (CR) където се инциира репликацията на мт ДНК. Репликацията на мт ДНК е асинхронна с клетъчното делене и често в цитоплазмата на всяка соматична или клетки от гаметогенезата може да има стотици или хиляди копия.
Митохондриалния геном на животните е много компактен, не е обременен с интрони или дълги интрагенни участъци на некодиращи последователности, характерни за нуклеарния геном. Тези генетични облекчения трябва да са силно селектирани при конкурентните цитоплазмени условия, когато по – малките мт ДНК молекули вероятно се реплицират по – бързо, отколкото по – големите и следователно притежават значително предимство при предаването в поколенията.
Предвид важната функционална роля на мт ДНК в животинската клетка и генералната структурна икономика на клетката, преди 1980 г. е имало спекулации с това, че този геном би трябвало да еволюира бавно в нуклеотидната си последователност. Изненадващо, това несе доказва на практика: както първо показват Wes Brown et al. (1979), животинската мт ДНК акумулира нуклеотидни субституции неколкократно по – бързо отколкото едноверижна нуклеарна ДНК. Няколко фактора вероятно водят до бързата еволюция на мтДНК: относително недостатъчни механизми за поправка на ДНК в митохондриите;оксигенните условия в митохондриите обусловени от наличието на среда богата на кислород; липсата на функционални ограничения поради факта, че мтДНК кодира само няколко типа белтъци и не продуцира протеини директно свързани с нейната репликация, транскрипция и транслация; факта, че мтДНК е „гола” не е свързана с хистонови белтъци, които сами по себе си са консервативни и могат да ограничават еволюционното ниво на нуклеарната ДНК. Бързата еволюция на мтДНК е също така отразена в сегашно време сред много от животинските видове, с висока вариабилност на нуклеотидната последователност,основа на филогеографските анализи.
Популациите на мтДНК молекули присъстват в соматичните и пре – полови клетки
Големи популации на мтДНК молекули /често хиляди и повече/ типично присъстват във всяка клетка, така потенциално усложняват генетичните анализи, нещо което не съществува за нуклеарните гени. /Всеки автозомен ген, присъства само в две копия на всяка диплоидна клетка и в едно копие на всяка полова/хаплоидна//. Въпреки това на практика повечето от индивидите са хомоплазмени по мтДНК, което означава че техните мтДНК молекули са принципно идентични по нуклеотидна последователност. Хетероплазмени животни /тези които показват вътреиндивидуално различие на мтДНК/ са регистрирани рядко, и в тези случаи молекулите се различават по една или няколко нововъзникнали мутации.
Защо хомоплазмията предоминира е неясно, но е по – вероятна възможността мтДНК молекули при пре – половите клетки да преминават през относително популационно стеснение, такова че големия брой на молекули при всяка зигота да са получени от по – малко молекули в пре – мейотичните пре – полови клетки. Така всяка генетична транзиция от хетероплазмия в хомоплазмия или от хомоплазмия в хетероплазмия може да се получи от генетичен дрифт в относително малко на брой генерации животни/ както и се оказва експериментално/. Друга теоретична възможност е тази, естествения отбор да е в полза на хомоплазмията, или механизма на генната конверсия периодично да хомогенизира мтДНК последователности в клетъчните линии. Независимо от тези случаи, хомоплазмията е важна за филогенетичните изследвания, понеже всяко индивидуално животно носи един, специфичен митохондриален генотип.
Друга отличителна характеристика на мтДНК е майчиното предаване на молекулата, тъй като цитоплазмата на зиготата основно произлиза от яйцеклетката. Всяко поколение получава мтДНК първоначално, освен ако не е изключение, от майка си, а не от бащата.
Женското предаване на мтДНК е подобно на мъжкото предаване на фамилните имена. В много човешки съобщества сина и дъщерята наследяват името на баща си, което само сина предава на поколенията. По същия начин синовете и дъщерите наследяват мтДНК от майките си, но само дъщерите ги предават на поколенията. Има няколко ключови различия: предаването на фамилните имена е условност приета от социума неотдавна /за пръв път се използва в Китай по време на Хан династията, приблизително в началото на новата ера/, докато предаването на мтДНК се случва от хиляди и милиони години; фамилните имена са характерни за хората, докато мтДНК „фамилни имена” присъстват природно при всички животински видове; освен това фамилните имена са съставени от няколко букви, докато мтДНК „фамилни имена” са съставени приблизително 17 000 нуклеотидни двойки /въпреки, че само част от тях се взема при типичните молекулярни изследвания/. Друга отличителна черта е, че всяко човешко фамилно име може да има няколко независими предшественици и така е полифилетично. Това се получава понеже източника на много фамилии е някаква професия /като Бейкър /пекар/ или Смит /ковач//, или видна историческа фигура / като Уилямс напр/, или превръщането на по – ранна система на патримония при която към името на даден човек се добява суфикс / Джонсон или Джаксон напр/. Обратно, всеки отделен мтДНК генотип принципно, с голяма сигурност е монофилетичен.
Понеже мтДНК се предава по единия родител, рядко, ако въобще някога рекомбинират. МтДНК се унаследява хаплоидно и безполово. Понеже при мтДНК няма пренареждането, което се случва по време на мейозата и сингамията, която се получава при сливането на половите клетки и засяга нуклеарната ДНК, мутациите които нармално определят генетичните вариации на мтДНК често се наблюдават в животинските популации. мтДНК генотипове се определят като хаплотипове, които варират помежду си с частични мутации натрупани и предадени от женския предшественик. Понеже мтДНК бързо еволюира, много и различни хаплотипове съществуват сред видовете.
Учените могат да използват тези хаплотипови последователности, за да определят майчината история на индивидите и популациите.
Генома на клетъчните органели има различни еволюционни правила при растенията.
Растенията имат два вида цитоплазмен геном – митохондриална ДНК /в митохондриите/ и хлоропастидна ДНК /в хлоропластите/ . Хлоропластите подобна на митохондриите също имат ендосимбионтен произход и също хлоропластидния геном е загубил част от гените си по време на еволюцията. Днес тези растителни органелни геноми са подобни на животинската мтДНК по своята структура /затворена циклична молекула/, начин на репликация /многобройни копия в клетката/, и не – Менделово унаследяване. Но също така те се различават от животинската мтДНК и помежду си по няколко важни молекулярни и еволюционни характеристики, както за пръв път е описал Palmer (1990).
Растителната мтДНК е много вариабилна по размер, варирайки от 20килобази до 2500 килобази при различните видове. Наследява се по майчин път, но не винаги така. Изненадващо растителната мтДНК еволюира бързо по отношение на генната подредба /генното реаранжиране е често срещано/, но по – бавно спрямо първичната нуклеотидна подредба. Така, еволюционната динамика на растителната и животниската мтДНК се различават диаметрално. Ниското ниво на еволюционна динамика на последователността на растителната мтДНК /около 100 по – бавна, отколкото на животинската/ плюс други технически усложнения означават, че тази мтДНК е слабо подходяща за филогеографски изследвания между видовете.
Растителната хлоропластна ДНК има други еволюционни правила. Тя слабо варира по размер между видовете /от 120 кб до 217 кб при фотосинтезиращите сухоземни растения/, като промяната на размера се дължи на удължаване на региона на дълга последователност от повтори. Молекулата има около 120 гени, които кодират рибозомни и транспортни РНК – и плюс някой полипептиди участващи в белтъчния синтез и фотосинтезата. Хлр(Хлоропластите)ДНК се предава по майчин път при повечето видове, от двамата родители при някой видове и по бащин път при други видове /повечето голосеменни/. Молекулата еволюира бавно, главно с генен реаранжимент, но също и с промяна на нуклеотидната последователност / около 3 – 4 пъти по – бързо отколкото растителната мтДНК, но много по – бавно от животинската/. Поради тези причини хлрДНК е доказано удобна за използване при оценка на филогенетични отношения при немалко таксони растения. С усърдно секвениране, достатъчно генетични вариации могат да бъдат разкрити, за филогеографски оценки между отделни видове.
Растителната хлрДНК принципно има много копия в клетъчната цитоплазма. Въпреки това повечето индивиди показват една хлрДНК хаплотипна последователност /и така са хомоплазмени/, а различните индивиди, между видовете често показват сериозни различия в последователността на хлрДНК. Така растителната хлр ДНК значително прилича на животинската мтДНК и учените могат по същия начин да използват нейния хаплоидна , нерекомбинираща природа, за да оценят генеалогичната история на растителните популации. Това проследяване може да бъде по майчина линия, по бащина линия или неспицифично и на двамата родители, в зависимост от начина на генетично предаване на хлр ДНК при отделните видове.
Нуклеарните локуси също имат генеалогична история
По принцип едно копие нуклеарни гени (scn) може също да осигури огромна последователност като основа за филогеографски изследвания на вътревидово ниво, но има три технически и биологически ограничения, които спират развитието на тези изследвания: относително ниското ниво на еволюция на последователностите на много от нуклеарните локуси; трудността да се изолират нуклеарни хаплотипове, от диплоиден организъм; феномена на вътрегенна рекомбинация.
Първото усложнение може да се превъзмогне с идентифициране на мониторингови scn последователности които еволюират много по – бързо спрямо останалите. Например интронните последователности на протеин кодиращите гени често еволюират по – бързо отколкото прилежащите им екзони. Втората пречка може да бъде преодоляна с използването на природната или изкуствена система на хаплотипната изолация. Например, гени на Х или Y- хромозомите на бозайниците /или Z и W – хромозоми на птиците/ са хаплоидни /хемизиготни в случая/ при всеки представител на хетерогаметен пол, както са всички scn локуси на мъжките от видове като пчелите или мравките с хаплодиплоидно полово определяне.. Когато гените в хаплоидно състояние не са осигурени природно /както са в случая на диплоидните организми/, е възможно да се пречисти нуклеарен хаплотип от всеки вид използвайки молекулярни методи /такива като изкуственото клониране в биологични вектори/. Независимо от използваните методи за генеалогични цели е важно да се изследват хаплотиповете индивидуално, защото иначе фазите на дадените хаплотипове /cis vs. Trans/ могат да останат неопределени при диплоидния индивид, което ще докаже хетерозиготността по много нуклеотидни позиции по продължение на гена.
Третата пречка заслужава уточнение защото недвусмислено е най –серозната. Във всяка популация на полово размножаващи се организми, мейозата и сливането на гаметите продължително пренарежда несвързани гени в нови мултилокусови комбинации. Тези рекомбинационни процеси могат да се случат и в рамките на даден локус, мокар че успешния мейотичен кросинговър се извършва между различни алели. Всеки рекомбиниран алел който се получава е като част на ДНК, омесена като амалгама, частите на която може да са имали напълно различно еволюционно минало. Така, вътрегенните /или вътреалелни/ рекомбинации се усложняват или представят неточно, което иначе казано е почистващ генеалогичен подпис от нуклеарния геном. Това контрастира с историческата чистота, характеризираща „ благословенно безбрачния” хаплотип на нерекомбиниращите цитоплазмени геноми (Dawkins, 1995). Въпреки това scn нуклеарни ДНК хаплотипове са използвани успешно при различни филогеографски изследвания (Hare, 2001), с едни от най – информативните резултати дошли от интрон последователности на протеин кодиращи гени. Понататък, дори и когато техническите и биологичните трудности на получаване на ясни генеалогични данни от scn нуклеарни ДНК последователности се окажат доказано непреодолими, не толкова директна информация за популационната история може все още да се добие от конвенционалните популационни генетични анализи, които се фокусират върху наблюдаването на честотата на филогенетични неподредени алели. Изследванията на алозими и микросателити, например, се използва рутинно за измерване на популационна алелна честота, което от своя страна може да бъде използвано при пространствено генетичните анализи на близкородствени популации. По специално когато такива данни са акумулирани на многобройни несвързани локуси, историческата информация често се случва да бъде сравнена с по – ясни генеалогични методи, които обикновенно се прилагат за цитоплазмените геноми.
Генеалогични концепции
Всички членове на популацията са свързани генеалогично
Въпреки многото различни генетични линии /аналогични на фамилните имена/, коитомогат да присъстват в популацията, те всички носят част от един общ предшественик , съществувал в някаква историческа точка. За да хванем тази концепция, разглеждаме първо майчината история на съществуващи индивиди при който е да е вид. Ако истинското популационно родословие е известно, всеки генеалог ще може да проследи майчините линии обратно във времето, и евентуално ще достигне до пра- пра – пра – пра – пра… пра бабата, няколко генерации назад, които са общи за всички сега съществуващи индивиди.Така всички сега съществуващи индивиди ще водят обратно в родословието до един общ предшественик по майчина линия. След това разглеждаме историята на съществуващата популация по бащина линия. Проследявайки обратно във времето по бащина линия иевентуално общия пра – пра – пра – пра…пра дядо ще бъде най – вероятно достигнат по подобен генеалогичен принцип. Важна особеност на майчината и бащина генеалогия, че те са като разклонено родословно дърво,/ не са мрежести/,въпреки че то е вградено в цялостното организмово родословие, което има твърде много разклонения вследствие от взаимното кръстосване. Всяка такава разклонена диаграма / събрани пътища, които са строго майчини или бащини/ , може да се счита като генно дърво или генетична генеалогия на организмовото родословие.
Трябва да се прави разлика между идеализираните генни генеалогии от една страна и не идеалните практически получени генни дървета от друга. Типична майчина генеалогия, например, е зададената по дефиниция действителна история на генетично предаване от майка на майка по генерациите, и типично бащино дърво е действителната история на генетично предаване от баща на баща на баща. Такива генеалогични истории не са познати директно от видовете в природата, но техните основни функции могат да бъдат оценени от подходяща ДНКпоследователност, сравнена между живи организми. Цитоплазмените геноми в частност могат да осигурят информация за майчината история, а Y хромозомата при бозайниците може да осигури информация за бащината история.
Клоните на генното дърво растат при всяка популация или видове
На пръв поглед може да изглежда така, че сумата от бащината и майчина история изчерпва всяка картина на генетично родословие. На това е далеч от истината на всяка популация на полово размножаващи се организми, защото повечето от нуклеарните гени са предадени и от двата пола. Представете си например факта ,че един индивид от неблизкородствено родословие има осем пра – пра родители и от всеки наследява приблзително 1/8от неговата автозомална ДНК,така само два от тези пра – пра родители / 25%/ ще бъдат индивидуални майчини и бащини прародители.
Принципно всяко организмово родословие може да бъде декомпозирано на многобройни генни генеалогии / фиг.1/ които са аналогни на майчиното и бащино гореописано родословно дърво, но което ще включва предаването по бащин /М/ и майчин /F/ предшественик /както е за всеки нуклеарен автозомален локус/. Например един хипотетичен път може да включва редуващи се полове / F, M,F,M,F,M,F,M…/ от предходните поколения, а друг може да бъде M, M, F, F, M, M, F, F…. При всяко родословие което е G генерации дълго, теоретичния брой на отделни „ генни дървета” от този вид /както е определено от полово – специфични генеалогии/ е просто 2G. В реалния свят няма автозомен ген, който да е предаден само по майчина линия или само по бащина линия, вместо това и двата пола са включени /макар и не точно определен полово специфичен начин по няколко поколения/ . При всяко събитие, на всяко поколение, броя на предаваните пътища възможни за алелите на отделен автозомален ген е увеличен четирикратно (M fi F, F fi M, M fi M и F fi F) спрямо мтДНК(F fi F само) или Y хромозомата(M fi M само). Това също предполага че времето на предаване по генерациите на автозомален ген е очаквано четирикратно по – назад в историята , всички други условия са еднакви, от тези на мтДНК и на Y хромозомата. Тази базова читрикратна разлика също може да също да се види като произтичаща от двукратния ефект получен от диплоидноста на автозомалните гени / за разлика от хаплоидната мтДНК/ съчетан с друг двукратен ефект, резултат от предаването от двата пола /за разлика от предаването по единия пол на мтДНК и Y – хромозомата/. По – общо казано , многочислени отделни нуклеарни генни генеалогии съществуват в родословието на всяка полово размножаваща се популация. С други думи несвързани сегменти на ДНК имат квази – независимо историческо предаване, така както техните генеалогични структури се унаследяват различно, най – малко до известно родословие, от локус на локус.
Фиг. 1 Хипотетично генно дърво или хаплотипно дърво /по– платните линии/ на 20 поколения организмово родословие. Квадратите са мъжките, кръгчетата – женските, а линиите свързватродителите с техните деца. Концепцията на генните дървета и генеалогичните процеси са основни за появата на филогеографията като отделна дисциплина на биогеографията.
Генеалогията и историческата демография са преплетени.
Родословното сортиране съпътства организмовата репродукция и прави този вид генни генеалогии,обсъждани по – горе. По отношение на майчината генеалогия, например, родословното сортиране няма да може да се осъществи само ако всяка женска във всяка генерация се унаследи само отедна дъщеря, при този случай няма да има йерахични майчини разклонения и предаване на наследствеността. Но в една реална популация, различни женски водят неравномерно до общия обем наследници, така че всяко майчино родословно дърво неизбежно се разклонява на няколко клони, наследниците се размножават или умират. Различните начини на размножаване поддържат неизбежния процес на предаване на поколенията.
За да илюстрираме това, да си представим че една женска роди дъщеря, съгласно статистически приетото разпределение на Поасон се означава /с отклоненията/ равно на 1.0. Поасон допуска че успешните дъщери, наследили майка си е независимо и случайно събитие. Съгласно този модел, очакванията майката да няма дъщери наследници в следващото поколение / или иначе казано очакването на майка без дъщери / е e=0.368 / където e е база на естествения логаритъм/ и нейната възможност за 1, 2, 3, 4 и 5 и повече дъщери е съответно 0.368, 0.184, 0.061, 0.015 и 0.004. Тези значения се добавят към едно поколение, но изчисленията / които са по – сложни/ се отнасят за много поколения. Те показват, например, че около 98% от всички основаващи майчини линии/матрилинии/ е вероятно да изчезнат след 100 поколения, дори и когато размера на популацията остава стабилен, а останалите 2% от матрилиниите оцеляват и продължават да се разможават.
Голям извод от такава статистическа теория е този, че исторически демографски фактори /такива като броя и вариациите на потомството на едно семейство/ може да се отрази на генната генеалогия по статистически придвидим начин. Например, точките на общите предшественици в генната генеалогия са по – стари в популациите с голям ефективен размер, при условие че всички други условия са равни. Така, практическите данни за дълбочината и разклоненията в структурата на генното дърво /като данни от мтДНК/ могат да се тълкуват съгласно генногенеалогичната теория , за да се изясни демографската история на популацията. Някой от прозренията на тази теория може да изглеждат нелогични. Например произхода от един предшественик в майчината генеалогия не предполага, че само една женска е живяла в това поколение, а просто показва че другите женски в това предшестващо поколение не са оцеляли до днешен ден с женски наследници. Друга важна точка е тази, че наследствието при майчината генеалогия не предполага, че другите женски в популацията са се провалили в предаването на генетичен материал на поколенията. Нуклеарните гени, от много от тези други индивиди могат добре да се представят в наследниците от алелите, които се предават не по майчин път в родословието. Аналогични твърдения и концепции могат да се направят и за бащините и всички други генеалогични пътища в рамките на разширяващото се животинско родословие.
Пространствено структурираните популации са генеалогично свързани
Филогеографските изследвания нормално се отнасят за многобройни популации разпространени по земята. Поведенчески и физически пречки на разпространение често са присъствали или присъстват в тези популации. Специално при тези видове с ограничена подвижност /възможност за придвижване/, популациите могат да бъдат частично изолирани една от друга на разтояние. Физическите филтри или бариери /такива като реки за сухеземните видове, или планини за равнинните видове/ често усилват ефекта на ограничената подвижност в развитието на популационната структура. Някой такива филтри могат да бъда полупропускливи и да позволяват случаен обмен на генетични линии, но други могат да блокират генния обмен въобще за дълъг период от еволюционно време. В последния случай, точката на общия предшественик за всяка генна генеалогия ще бъде не по – малко стара от появата на тази бариера. Целите на филогеографските оценки е да се използва генното дърво, за да се покажат историческите и съвременни сили, които образуват сегашната генеалогична архиктектура на популациите и близкородствените видове.
За всеки вид който генетично се разделя от някаква преграда на разпространението му, генеалогичния начин зависи от продължителността на действие на тази бариера, както и от историческата демография на разделените популации. Да разгледаме например , случайно чифтосващи се видове от предшествена популация, които се разделят на две отделни популации (A и B) от някакво изолиращо събитие. Първоначално е най – вероятно популациа А и В да са полифилетични спрямо майчиния произход /или мтДНК отношения/, което означава, че някой индивиди от А са генеалогично по – близко до някой индивиди от В, отколкото с други индивиди от А /или обратно/. След няколко поколения, при сортирането на отделните линии, в крайна сметка може да се получи така, че популация А да стане парафилетична спрямо В /или обратно/, което означава, че някой индивиди от А ще са генеалогично по – близко до някой индивиди от В, спрямо други индивиди от А, но всички индивиди от В са станали монофилетична единица по време на комбинирана генеалогия на популациите А и В. След още няколко поколения, А и В евентуално могат да станат реципрочно монофилетични по отношение на майчиния предшественик, което означава всички индивиди от съответната популация са едни на друг близки и генеалогично родствени по този специфичен компонент от родословието. Тази генеалогична транзиция от полифилия през парафилия до реципрочна монофилия в генното дърво е очаквана от генногенеалогичната теория, всеки път когато смесени предшествени популации се разделят на изолирани еденици /фазата на полифилия може да се пропусне, ако предшествената популация вече е била разделена преди това от някаква бариера/. Времева продължителност на транцизионните генеалогични етапи също зависят от големината на дъщерната популация, защото целия генеалогичен процес се повлиява от сортировката на линиите или реорганизацията на линиите произтича от диференциалната репродукция. Важна концепция е тази, че даден генеалогичен начин сам по себе си може да бъде еволюционно динамичен /по – скоро от статичен/ последван от събитие на разделяне на популацията.
Генеалогичните съответствия са доказателство за дълбочината на филогеографската структура
Предвид очакваната хетерогенност но генеалогичните пътища между видовете и многообразието на поведенчески, екологични и еволюционни сили които оформят генното дърво, как може филогеографските истории на популациите да бъдат реконструирани посредством молекулярни данни с голяма достоверност? Специфично, как може исторически дълбоки /древни/популационно генетични подразделения да бъдат емпирично отличени от тези които са еволюционно плитки /скорошни/? Отговорите на тези въпроси се намират в принципите на генеалогичното съответствие (Avise & Ball, 1990).
По принцип, най – малко четири различни аспекта на генеалогичното съответствие или съгласуването сред белезите може да се определят.
Аспект I съответствие е в генния локус: ако много нуклеотидни субституции на последователността на ДНК твърдо отличават един хаплотип от друг, дълбоко /противоположен на плитък/ историческо разделяне може сигурно да се добави в разклоненията на генното дърво при изследванията. Това е истина понеже акумулирането на нови мутации е бавен процес, затова значително време трябва да е минало преди даден нерекомбиниращ ДНК хаплотип да покаже големи разлики в последователностите. Ако генеалогичното разделяне е документирано само за едно генно дърво, разделянето в това отделно дърво е можело да стане и по други причини /такива като балансиращия отбор, който може да е в полза на продължително възпиране на крайно отличаващи се алели на даден локус/.
Аспект II съответствие е в мулти – локусов план: ако дълбоко разцепване на много генни дървета твърдо отличава същия набор от популации, еволюционните сили отговорни за това, трябва да са имали значително геномно влияние. Най – походящия кандидат за такъв широкобхватен ефект на локуси е продължителното спиране на генния поток, такъв който възниква от продължителни физически, поведенчески пречки на разпространение. Така всяко емпирично документиране на съответствие в Аспект II, твърдо предполага дългосрочно историческо разделяне на популационно ниво.
Аспект III съответствие е в мулти видов план: ако пространствено последователно генеалогично разцепване се показва сред няколко близко разпространени видове, отговорните за това еволюционни сили трябва да са били със широко разпространен ефект на ниво биотична система или екосистема. Накрая, аспект IV съответствие се отнася за всяко м съответствие между няколко линии на емпирични доказателства. Например, ако генното дърво се резцепва при отделни видове и географски е следвано от продължителни препятствия в разпространението , които се знаят или се предполагат от независимо доказателство / морфология, подвидова таксономия или физическа геология/ дълбоко филогеографско разцепване между видовете твърдо може да се предположи. Някой от генногенеалогичните концепции, разглеждани в този раздел, може да изглеждат езотерични. Въпреки това те всички са заимствани от живия свят с интерпретация на практическите филогеографски модели, както ще покажа в следващия раздел.
Практически филогеографски модели
Здрава генеалогична стурктура характеризира слабо разпространените видове
Първите основни филогеографски изследвания на мтДНК включват югоизточния джобен гофер, доста седентарен копаещ гризач (Avise et al., 1979). В генетично изследване на 87 животни, разпространени в Джорджия, Алабама и Флорида, са определени повече от 20 различни мтДНК хаплотипове. Някой от тях се различават само по един или два изследвани нуклиотидни позиции, докато други имат по – големи различия. Всички тези мтДНК хаплотипове са доказано пространствено локализирани и близко свързаните хаплотипове нормално са географски близки. В добавка, дълбоко генеалогично разцепване отличава всички гофери от източната от тези от западната част на видовия ареал. Плитките разцепвания на генното дърво са показателни за рутинна изолация на разстояние, докато дълбоките разцепвания /изчисления на 3% нуклеотидна разлика/ вероятно регистрират по – дълга история на генетично разделение – може би повече от милион години от своя общ женски предшественик. Така ранни практически изследвания маркират раждането на филогеографията като отличаваща се дисциплина.
През последните 30 години, подобни филогеографски оценки, базирани на мтДНК последователности са проведени буквално стотици на слабо подвижни животински видове, различни малки бозайници, влечуги, земноводни до безгръбначни като сухоземни охлюви и щурци. За толкова разнородни организми, филогеографските резултати са типични като тези по- горе описани: за всеки вид, близки, подобни или идентични мтДНК хаплотипове са географски локализирани и малък брой /типично от 1 до 6/ дълбоки генеалогични разцепвания, обикновенно отделят регионалните популации, които вероятно се диференцират в отделни рефугии (Avise, 2000). По дълбоките разцепвания обикновенно се свързват с известно или вероятно препятствие за разпространение, такива като Панамския провлак, който се образува преди около 3 млн години и разделя много тропични морски видове /риби, бодлокожи и др/ на Атлантическа и Тихоокеанска популация, които сега са доста различаващи се помежду си.
Сходни филогеографски оценки, базирани на хлр ДНК са направени на няколко растителни вида. Например, между 85 популации на обикновенния бук генетично са изследвани в рамките на ареала на разпростаренение на вида в континетална Европа. 11 ралзични хлр ДНК хаплотипа са отличени (Demesure et al., 1996). Някой от тези хаплотипове са различаващи се по една или две позиции, докато други са различаващи се по повече. Главните открития са, че един хлр ДНК хаплотип предоминира в по- голямата част от Европа, докато други близкородствени на него са близко географски локализирани и един крайно различаващ се набор от хаплотипове има в Крим. По – широко разпространения хаплотип е вероятно последователността на предшественика от западна Европа /от който другите билзкородствени хаплотипове са възникнали с по – скорошна мутация/, а отличаващите се хаплотипове от Крим вероятно показват наличието на втори плейстоценски рефугиум за този вид в източна Европа.
Подвижните създания също могат да бъдат генеалогично структурирани
Много организми имат голям потенциал на разпространение, като възрастни форми /например крилатите насекоми, или рибите на откритите води/, като ювенилни форми /например морските безгръбначни с дълго живееща пелагична ларва/, или като гамети /например растителните видове с полен, разпространяващ се от вятъра/ Дори и при тези подвижни видове, исторически бариери на разпространение могат понякога да бъдат непреодолими. Например Панамския провлак блокира вътреокеанския генен поток на почти всички морски видове след появата си преди 3 млн.г. Това е рефлектирало в съвремието при голям брой от морските риби, скариди, морски костенурки и други видове, на които популациите от Атлантика и от Тихия океан са различни от двете страни на провлака. Те показват доста дълбоко историческо разделение при молекулярни изследвания на мтДНК и други гени (Knowlton et al., 1993; Bermingham et al., 1997).
Понякога дълбоко генетично разцепване разделя съществуващи популации, които сега не показват да са разделени от някакви пречки в разпространението. Типичен пример е
морската лещанка (Fundulus heteroclitus), която обитава амерканското атлантическо крайбрежие, където е регистрирано голямо генеалогично различие между северната и южната популация на основа на мтДНК и секвениране на нуклеарни гени, кодиращи лактат дехидрогеназа (Bernardi et al., 1993). Този пример на аспект II съответствие може да е доказателство на присъствие на видовете в два отделни изолирани рефугиума по време на плейстоцена където генетичната диференциация се натрупва, последвано от разширяване на ареала и вторичен контакт. Не може да не се каже, че естествения отбор също не оказва влияние за получаване на този краен филогеографски резултат. Например, естествения отбор в миналото може да развие някой от генетичните различия, които да останат в алопатрията, и обратно естествения отбор /от факторите на околната среда като температура на водата/ могат да инхибират хаплотипния обмен между северния и южния региони. Подобен пример включващ аспект II на съответствие, и за който подобни обяснения могат да се направят се отнася за ракообразното Tigriopus californicus, разпространено покрай американското тихоокеанско крайбрежие (Burton & Lee, 1994).
Някой организми с голям потенциал на разпространение могат да покажат очевидна популационна генетична структура, резултат по скоро от филопатрия /поведенческа вярност към определени места/ отколкото от физически бариери сами по себе си. Няколко китоподобни видове, например, матрифокални /социално организирани около женските/, и имат очакван генетичен почерк, който е матрилинеална популационна генетична структура. При мтДНК изследвания на гърбатите китове (Megaptera novaeangliae), популациите на всеки от трите океана са доказано подразделени по майчини кланове, които вероятно възникват и се поддържат от женската привързаност към отделни миграционни пътища. (Baker et al., 1993). Тази привързаност вероятно се развива, още когато малкото следва майка си в пътя на миграциите и, от водите на високи към ниски географски ширини. Мигриращите птици са също така силно подвижни, но няколкото изследвани вида показват значителна филогеографска структура на основа на гнезденето /а също така понякога и по миграционните пътища и местата на зимуване /. При тъмногръдия брегобегач (Calidris alpina), например, пет мтДНК клона отговарят на регионалните популации на гнездене в Европа, източен Сибир, централен Сибир, Аляска и Канада (Wenink et al., 1996). Този вид на популационна генетична архиктектура при мигрантите на дълги дистанции е очевиден резултат на верността на поколенията към отделните миграционни пътища.
Не всички видове са филогенетично подразделени
Въпреки че доста видове показват значителна филогеогорафска структура, съществуват изключения, и те могат да бъдат доста информативни за ширината на филогеографските принципи. Един вид, който видимо няма очевидно филогеографско разделение е червенокрилия дрозд (Agelaius phoeniceus), един от най – популярните птици в Северна Америка. В генетични изследвания на 127 индивида от целия континент, са определени 34 различни мтДНК хаплотипа,един от тях е най – срещащия се /практически повсеместен/ , доказано в сърцевината на „ звездоборазуването” на генеалогията, към който всички останали хаплотипове /всеки сам по себе си рядък/ са независимо свързани (Ball et al., 1988). Такъв „звездообразуващ” тип е очакван почерк за видове, които са се разпространили в последно време от един географски източник; най – често срещания и най – повсеместнен хаплотип е вероятно този на предшественика, от който по – редките хаплотипове са възникнали чрез отделни мутации. Така генеалогичните данни от мтДНК предполагат че, следвайки отеглянето на ледниците / започнало преди 18 000 г./, червенокрилия дрозд разширява широко неговия ареал и размера на популацията си, като колонизира континента от първоначален Плейстоценски рефугиум.
Друг пример от този основен тип е показан от американската змиорка (Anguilla rostrata). Този вид е катадромен, което означава че индивидите прекарват по- голямата част от живота си в реки и езера, но евентуално мигрират към морето, за да хвърлят хайвера си и умират /за разлика от анадромните видове като сьомгата, която живее в морето, но мигрира в реките, за да хвърли хайвера си/. Миграциите за хвърляне на хайвера от емериканските змиорки в Саргасово море, в западната тропична среда на Атлантическия океан, където мигрантите от източна Северна Америка се събират за случайни чифтосвания – панмиктични. Получените ларви после евентуално поемат по своя път обратно в пресноводните хабитати. Дали този странен жизнен цикъл означава, че ювенилните змиорки от техния широкообхватен пресноводен ареал са част от един добре смесен генофонд? Филогеографските данни получени от мтДНК и нуклеарни гени са напълно в съответсвие с тази възможност. Хаплотиповете и алелите са повсеместно разпределени /и във близки честоти/ сред изследваната популация на змиорките от Мейн до Луизиана(Avise et al., 1986).
За всеки такъв вид, които има един, добре смесен генофонд, генногенеалогичната теория, може да се прилага към генеалогичните данни, за да се оцени дали вида има дълговременен ефективен популационен размер. Ефективния брой на индивидите (Ne) се отнася за размера на идеализирана популация, която има същите генетични свойства /същото време на общо развитие/ като това, наблюдавано в реалната популация. Обикновенно, дългосрочно или еволюционно Ne е доста по – малко от N /размера на преброеното/ по една или няколко следни причини: броя на чифтосващи се мъжки може да варира от броя на чифтосващи се женски; популациите в природата обикновенно варират по размер във времето, и могат да преминат през известни стеснения; някои индивиди могат да оставят повече потомци от други, създавайки здравословно разнообразие в семействата; могат да се случат селективни помитания при които позитивно отбрани мутационни помитания се случват в популациите или видовете, като така свиват предшествениците до по – малко нидивиди, отколкото може да има; или други фактори, такива като периодичните изчезвания или реколонизации, които могат да доведат видовете да покзват по – ниско Ne отколкото могат да се предвидят от наличното преброяване. Генногенеалогичния подход за изчисляване на еволюционното Ne, включва оценка по молекулярни данни, хистограма на еволюционното време за поделения предшественик на всички двойки изследвани индивиди, и последващо сравнение на „несъотвестващото разпределение“ на наследствените очаквания /според неутралната теория/ за хипотетични популации с различни ефективни размери. За змиорките, чифтово изчислено по мтДНК последователности, разклоняване се превръща на времето за развитие на всички поколения като се има предвид, че продължителност на всяко поколение е 10 години и че тяхната мт ДНК еволюира със стандартна стъпка /2% секвенционна разлика на млн години/. Резултантното несъчетаващо се разпределение доказва, че е близко по сила и вид на очакванията за общото развитие на поколенията за една популация с еволюционно ефективен размер Ne = 5,500 женски (Avise et al., 1988). Качествено подобни изводи са направени за няколко други вида, които показват генетично добро смесване. Типично /както при американските змиорки/, еволюционно ефективните популационни размери са в един порядък по – малки, отколкото реално преброените популационни размери, може би защото всеки вид е минал през популационни стеснявания, един или няколко пъти в своята еволюция и/или поради други демографски фактори /упоменати по – горе/, които могат да намалят дългосрочно ефективния популационен размер под нормалните нива на преброяването.
Историчните процеси също се отразяват на биотичните съобщества
Няколко регионални, за дадена природа, генетични анализи сравняват филогенетичните изводи сред много близко разпространени видове (Bermingham & Moritz, 1998). Забележителни,но последователни открития са тези, че видовете с подобни ареали или екологии, често са генеалогично структурирани по подобен начин.
Това предполага, че исторически биогеографски процеси са не само видово – идиосинкратни по техния ефект, а вместо това имат също така съгласувано оформяне на генетичните архиктектури на дадените регионални биоти.
Един пример на аспект III на генеалогично съответствие е осигурено от генетични изследвания на крайбрежна фауна на югоизточните САЩ, където популации от Мексиканския залив при всеки от няколко вида са силно генетично различаващи се от доста популации по Атлантическото крайбрежие (Avise, 1992). Дълбоки генетични разделения между Атлантическите и тези от Мексиканския залив популации не са универсални при всички изследвани видове /американската змиорка дава точен пример/, но по способ на подреждане предполагат, че значителна част от крайбрежната биота е била генетично повлияна по един общ съгласуван начин. Филогеографските фактори отговорни за този случай не са напълно изяснени, но работна хипотеза е тази, че предшествени популации в Мексиканския залив и в Атлантика са исторически разделени, може би по време на Плейстоцена, и са натрупали генетични различия, които днес се поддържат от екологични препятствия /такива като океанските течения или разлики в температурата на водата/ за междурегионален геннен поток. Съгласувани генеалогични способи, за които историческите обяснения са по-ясни, включват флората и фауната в Европа. Генетични изследвания осигуряват сигурни доказателства, че три Европейски региони не са били покрити от ледници по време на Плейстоцена – Иберийския, Апенинския и Балканския полуострови – и са били първоначални рефугиуми за много животински и растителни видове, които в към днешно време реколонизират континента (Hewitt, 2000; Weiss & Ferrand, 2006). Филогеографски отпечатъци са очевидни днес при пространствения аранжимент сред много видове от различни генеалогични клонове /често наричани вътревидови „филогрупи”/, които ни връщат обратно през различни колонизационни пътища към един или два от тези плейстоценски рефугиуми. В много случаи, пост плейстоценския район на разширяване също води до такива отличаващи се филогрупи във вторичен контакт по границите на групите, където може да има хибридизация. Тези гранични зони са концентрирани в специфични ареали /отбелязани в Алпите, централна Европа и Скандинавия/, може би защото ледниковите рефугиуми са общи, историите на постглациална експанзия и на физически пречки на разпространение също.
Историята понякога се повтаря
Друга илюстрация на всичките четири аспекта на генеалогичното съответствие включва рибите от югоизточните САЩ. За всеки от няколкото вида изследвани по мтДНК , а някои и по нуклеарните гени, популациите в западните реки /които се отвеждат в Мексиканския залив/ са генетично различни от популациите в повечето източни реки /отвеждащи се в Атлантическия океан/ и полуостров Флорида (Avise, 2000). Подобни гранични зони също съществуват там, където два форми се припокриват и хибридизират. Тези открития твърдо предполагат присъствието на два отделни рефугиума, в които генетичните различия са се натрупали и от които е настъпилопоследващо разпространение. Въпреки, че тези филогеографски начини са генерално съгласувани сред много таксони, дълбочината на генеалогичното разцепления се различава значително от вид на вид. Една възможност е просто, че молекулярните еволюционни степени се различават при тези таксони, но е по интригуваща хипотезата че, тези различни съвременни видове може да са били исторически повлияни по различно време. През 2-те млн год на плейстоцена, контитенталните ледници са се образували и изчезвали циклично и така са ограничавали и освобождавали отделните рефугиуми. Може би някои от съвременните популации риби в източните и от западните водоеми водят обратно към един и същ рефугиум, но по различно време от глациалните цикли. За това ще е необходимо в един от рефугиумите по време на глациалните цикли предшествена популация да е изчезнала и да е заменена по-късно от колонизатори от друга. Мрежестия резултат би бил съгласуван в пространствен аранжимент на генното дърво, но с различна времева дълбочина на техните разцепвания. Тази вероятност, въпреки че е силно спекулативна, е също подкрепена от различни типове от биогеографични доказателства: структура на съобществата. Няколко видове риби в югоизточните/Атлантически/ или западните /Мексикански залив/ реки, предполагат че популациите някога обитавали една или друга от тези ареали са изчезнали /или може би са дивергирали толкова много, че им е даден нов таксономичен статус – вид/. Ако е така, това ще помогне да се обясни не само общите фаунистични модели, споменати по – горе, но също така защо тези две биогеографични провинции са принципно съгласувани с две главни молекулярни филогрупи, съществуващи сред отделните видове.
Дори и митохондриалното генно дърво само по себе си може да даде ключови биологични прозрения
Въпреки, че филогеографските анализи най – добре се базират на нуклеарни и на митохондриални генни данни, мтДНК генеалогия сама по себе си може да е достатъчна за някой цели поради своята специална връзка с женската популационна демография (Avise, 1995). Да си представим, например, един вид при който женските са седентарни, а мъжките се придвижват на големи разстояния и се чифтосват на случаен принцип с женски от различни места. Матрилиниите при тези видове ще бъдат точно структурирани, а нуклеарната генеалогия ще бъде географски разпределена на случаен принцип. Въпреки това крещящо разногласие между мтДНК и нуклеарните гени дърво, матрилинеарната структура сама по себе си означава, че всяка географска локация е автономна от гледна точка на размножителния резултат и това от друга страна може да има сериозни последствия за управлението на популацията и нейната консервация. За да се аргументираме по – конкретно, да разгледаме зелената морска костенурка (Chelonia mydas). Тези костенурки се събират по различни места от световните тропични океани и излизат на брега, за да снасят яйца. Иначе прекарват по – голямата част от живота си в морето, хранейки се на места, които могат да бъдат отдалечени на стотици и хиляди километри от местата за полагане на яйцата и така извършват Херкулесови репродуктивни миграции. Генетичните изследвания показват, че групите за полагане на яйца се различават едни от други по мтДНК хаплотипове (Bowen & Avise, 1996), което означава, че женските имат привързаност към местата за полагане на яйца /защото ако нямат, мтДНК хаплотипове трябваше да са смесени в различните места за полагане на яйца/. Така, ако костенурките от всички места за полагане на яйца се чифтосват на случаен принцип и техните нуклеарни геноми са измесени като резултат от това случайно чифтосване /а практически те не са/, превързаността на женските към местата за полагане на яйца, документирано от мтДНК ще показва значителна степен на демографска автономност за всяко място. Този извод има консервационно значение: ако на някое от местата за полагане на яйца се унищожи от природата или от човешка дейност, установяването на ново място на това място за гнездене е малко вероятно и още повече за кратко време. /Естествено, изоставянето на някое от тези места също се случва, но затова видът има много места за полагане на яйца/. Възниква и друг интересен контекст на матрилинеарната история сама по себе си. Зелените костенурки ни дават такъв пример. Едно от главните места за полагане на яйца на този вид е остров Асенсион, един малък скалист вулканичен остров намиращ се посредата между Южна Америка и Африка. Всеки сезон, женските, които иначе се хранят край Бразилското крайбрежие се отправят в едно 4 400 километрова одисея за полагане на яйца до остров Асенсион и обратно. Защо предприемат такова пътешествие, което изисква невероятни физиологични усилия и навигационни умения, когато има близки места за полагане на яйца, намиращи се по крайбрежията на Южна Америка? Една интригуваща възможност предположена преди години от Архи Кар /първия биолог, изучавал тези костенурки/ е че тези костенурки са започнали да полагат яйца на този остров още когато този остров е прото – остров преди 80 млн год.. Тогава Южна Америка и Африка са били разделени от много тънка морска ивица. През изминаващите хилядолетия континентите бавно са се раздалечавали от тектоничните движения, местата за снасяне на яйца на женските постепенно са се отдалечавали при всеки миграционен цикъл със сантиметри на поколение. Тази хипотеза, ако е истина, трябва да бъде потвръдена от ненарушена матрилиния от поколения предшественици прези окло 80 млн год до сега, които са гнездяли всички на остров Асенсион, и също така предполага голямо различие на мтДНК последователности акумулирани между тези костенурки, снасящи яйца на Асенсион и другите, снасящи айца в Южна Америка или другаде. Хипотезата на Кар е подложена на практическо тестуване и резултатите са фалшифицирани (Bowen et al., 1989). мтДНК хаплотипове на женските костенурки от Асенсион се различават от тези полагащи яйца в Южна Америка, но степента на различие на последователностите е много малка, показваща че матрилиниите на костенурките, полагащи яйца сега на Асенсион ни връщат към предшественик колонизатор много по – ранен, преди около 100 000 години най – много.
Преспективи на филогеографията
През последните три десетилетия филогеографските преспективи /митохондриалната и на други гени дървовидна система от които тези преспективи възникват/ добавят много неортодоксални прозрения за еволюционните генетични процеси (Avise,2007). Таблица 1 кратко сумира 21 такива концептуални и емпирични приноса. Филогеографията като дисциплина се развива фантастично през последните 30 години и не показва признаци на забавяне. Напротив, аз виждам възможности за разширение по най – малко три припокриващи се фронта.
Таблица 1. Много неортодоксални преспективи на еволюцията са възникнали от молекулярни открития на мтДНК и чрез филогеографски преспективи, мотивирани от тези открития. За пояснение, вижте долния текст и също така Avise (2000, 2007).
Популационна йерархия. Всички популации на мтДНК молекули присъстват в соматичните и полови клетъчни линии и така добавят едно изцяло ново ниво на популационно генетичната йерархия.
Стеснения при половите клетъчни линии. Хомоплазмията типична за животинската мтДНК, предполага че при популационни стеснения в половоклетъчните линии вероятно ще се предадат митохондриално наследствено.
Консервативна функция и бърза еволюция. Бързата еволюция на животинската мтДНК е предизвикателство за идеята, че функционалните ограничения задължително налагат ниска скорот и ограничават промените в последователностите на ДНК.
Асексуално предаване. Цитоплазмените геноми при видовете с полово размножаване нормално показват клонално / еднородителско, нерекомбиниращо/ генетично унаследяване.
Матрилинеарни истории. Бързото ниво на мтДНК еволюция, съчетано с майчино унаследяване на молекулата, означава че могат да се възстановят вътревидовите майчини истории.
Индивидите като оперативни таксономични единици. Понеже всяко животно има специфичен мтДНК хаплотип, всеки индивид може да се разглежда като една „оперативна таксономична единица” за генеалогични оценки.
Концепция за генното дърво. При организмите с полово размножаване, мтДНК осигурява прототип пример за неанастомозираща или не – ретикуларна, немрежеста генна генеалогия, или” генно дърво”.
Разширена концепция за генното дърво. По принцип, мултигенерационно родословие на всеки вид, размножаващ се по полов път съдържа в нуклеарния геном, многобройни неизоморфни генни дървета.
Генни дървета срещу видови дървета. Генните дървета за несвързани нуклеарни локуси могат и често се различават топологично от съставна видова филогения; несъответствията между видовите и генните дървета, дължащи се на идиосинкратично сортиране на линиите, могат също да характеризират съществуващите таксони които са се отделили в миналото.
Генеалогични топологии. Във връзка с дадени генни дървета, филогенетичния статус на всяка двойка родствени видове е само по себе си еволюционно динамично, с общ времеви ход на събитията, следващо за популацията или вида разделение, което да е полифилия, парафилия или реципрочна монофилия.
Генеалогично съответствие. Поради идиосинкратичната природа на индивидуалните генни дървета, дълбоки разцепвания на популационно или видово ниво могат да се разпознаят само във връзка с някой от различните аспекти на „генеалогичното съответствие” /наличие на един и същ белег при двата алела , сред локуса, сред вида, или по различни линии на доказателства/
Генеалогия и демография. Сред някои видове, процесите на развитие описват връзките между генеалогия и популационна демография и показват , че генеалогия и демография за са внимателно преплетени концепции.
Генеалогия и пол. Ключови демографски и поведенчески параметри често се различават между женските и мъжките по начина на отразяване на полово – специфичните генеалогични структури.
Ефективен популационен размер. Емпирични генно – дървесни изследвания, обяснени чрез теорията за съвместното развитие, показват че повечето видове имат изненадващо малки еволюционно ефективни популационни размери.
Вътревидови популации. Емпирични данни показват, че отделнити видове често са подразделени на ясно разграничени генеалогични единици /вътревидови филогрупи/.
Брой филогрупи. Емпирично, общия брой на ясно разграничените вътревидови филогрупи на вид обикновенно са малко (c. 1–6).
Пространствена ориентация.Вътревидовите филогрупи най –често алопатрични и обикновенно са пространствено ориентирани по начин, които видимо е с добър биогеографски замисъл.
История на природата. Генеалогичните данни показват това, че допълнително към историческите биогеографски събития, поведение и историята на природата на видовете също се отразява на филогеографските модели.
Таксономия и консервация. Филогеографските данни и концепции са много уместни за таксономични решения и също така за консервационната биология.
Филогенетични доводи. По принцип, и противно на конвенционалния разум, езика и концепциите на филогенията /генеалогията/ са силно свързани на вътревидово ниво.
Микроеволюцията, подобно на макроеволюцията има история. По принцип, вътревидовата еволюция рядко образува вид равновесни резултати /такива като тези получени при островния модел на популационна структура/ които да са математично обяснени, обект на традиционната популационна генетична теория; вместо това вътревидовите генетични архиктектури са наследени исторически /генеалогично/ и идиосинкратично по биографичен контекст. Полезрението на филогеографията ясно разпознава и побира тази фундаментална реалност.
Разширяване на емпиричните анализи на многобройни нуклеарни локуси
Много филогеографски анализи досега са били фокусирани първоначално на цитоплазмените геноми /поради няколко логически защитени причини, дискутирани по – рано/. Получената информация от тези относително лесно получени резултати бе голяма, но в търсенето на нови средства, един значителен напредък в тази област носят генеалогичните резултати от изследването на потенциално интересния, но трудно достижим нуклеарен геном. Преодоляването на техническите трудности /такива като бързо изолиране на нуклеарни хаплотипове от диплоидни индивиди/ и естествените пречки /по – специално как да се справим с вътрегенната рекомбинация/ може да е обезсърчително, но всеки изследовател, успешен в тези начинания, веднага би станал научен пионер. Целите са, да се възстанови генната генеалогия от многобройни несвързани нуклеарни локуси с идеята да се търси модел на вътре – локусово генеалогично съответсвие / или липсата му/, което ще носи сериозни интрепретации на историчните записи на всеки вид.
Разширяване на генногенеалогичната теория
Генно генеологичната теория е възникнала първоначално с ориентация към един локус, така съществува една голяма стая за адресиране на вътрелокусови вариации в генногенеалогичните резултати като функция на различни генетични фактори /такива като степента на свързаност между локусите/ и за миналото и бъдещето на популационно демографските параметри. Значителна мултилокусова генногенеалогична теория също ще трябва да се съгласува към много варианти на пространствени популационни архиктектури, които природата може потенциално да сътвори. Свързана с тези усилия ще е необходимостта към по нататъшно интегриране /свързване/на очакванията от генногенеалогичната теория и други аналитични методи (Excoffier, 2004) с емпиричните данни в по тясно свързани, интелигентни комплекси. Целта на комплексите трябва да бъде твърда „статистическа филогеография (Knowles & Maddison, 2002; Templeton,2004; Knowles, 2008), която да отразява реалистична интеграция на популационната генетика и филогенетичните методи.
Разширяване на филогеографските анализи към други таксони
Въпреки, честотици от практическите изследвания във филогеографията са публикувани, само една малка част от биологичния свят е изследвана по отношение на филогеографски преспекиви. Така че бъдещите усилия трябва да разширят филогеографските оценки към много други таксони. Доста от тази работа би била мотивирана от желанието да се разберат идиосинкратичните генеалогични истории на отделните видове, което може да е от специален интерес поради таксономичните неточности, консервационни отношения, очарователни природни истории или от други биологични мотивации, които да са видово специфични. Главния изследователски фокус също така може да е поставен върху сравнителни филогеографски оценки на мулти видови регионални биоти. Основани на минал опит /такъв като с различни таксони в югоизточните САЩ и в Европа/, поделени филогеографски модели /съгласуваности/ рефлектиращи на видимите исторически събития може би ще бъдат доказани вхарактеризирането на много видове от много регионални фауни и флори. Наистина, колаборативни усилия са вече напът да разкрият филогеографски модели на няколко континента и при различни субконтинентални региони, като целите им са да реконструират главните насоки на скорошновремевитеистории на изолиране и разпространение във всеки един ъгъл на света.
Мястото на филогеографията сред науките за биоразнообразие
Преди началото на „филогеографската революция” в края на 1970г и на 1980г, относително малко връзки са правени между биолозите изучаващи вътревидовите /микроеволюционни/ генетични процеси и тези анализиращи надвидовите /макроеволюционни/ генетични модели. Досега тази област беше на популационните генетици., които се занимават с промените в популационните алелни честоти, произлели чрез мутации, генетичен дрифт, генен поток, естествен отбор и полов отбор. В последно време с това се занимават филогенетиците и систематиците, които работят най – вече с еволюционните отношения на видове и надвидови таксони. Основни дисциплини с които работи популационния генетик са статистиката, математиката, екологията и популационната демография, и вероятно с малко от филогегорафските принципи. Работата на систематика е свързана с отделни таксономични групи в музея или на полето, но вероятно с малко от принципите на популационата генетика. Това положение на отношения е и жалко и иронично, защото популационната генетика и филогенетичната биология трябва да са част от времевия еволюционен континуум. Понеже филогеографията се отнася до изследвания на близкородствени организми и свързани видове, тя би трябвало да се подразбира като клон от популационната генетика по своя специален фокус върху генеалогията /не само върху алелните честоти/ както и се вижда от историческите генетични записи в отделни части на ДНК. Най – точно казано, вътревидовата генеалогия може да се интрепретира като филогения, разглеждана по отблизо. Понеже филогеографията си взаимодейства с генеалогията / микрофилогения/ тя също може да се разглежда като клон и на филогенетичната биология. Но се отличава от традиционната филогенетика по своя специален фокус върху популационната история и демография. Традиционно настроение сред систематиците е, че филогенията няма значение на вътревидово ниво, понеже генетичните линии са гъсто преплетени и анастомозиращи при едни размножаващи се по между си популации. Както видяхме, региони от свързани нуклеотиди/ мтДНК или хлрДНК са само два примера/ имат дървовидна история сред видовете и разклонените структури на тези генни генеалогии носят значителна аналогия / макар и често в различен времеви мащаб/ на филогенетичните дървета сумирайки историческите взаимоотношения сред репродуктивно изолирани видове и таксони. Освен това, систематиците рядко разглеждат и имат предвид историческата популационна демография при реконструирането на видовите филогении, а наистина историческите демографии на родствени организми имат важно и продължително влияние на начина на наследствените вериги, свързващи популациите и видовете по време на еволюционния процес. Филогеографията и генногенеалогичната теория биха били изяснителни отколкото генеалогията и историческата популационна демография са, взаимно свързани. Накрая, филогеографията работи с пространствените и времеви параметри на генеалогията и така може да бъде разглеждана като клон на биогеографията /може би нейния най – естествен дом/. Тя отново се различава от традиционната биогеография по своя специален фокус на родствените популации и на подрабната генеалогична информация. Тя също се различава от екогеографията/ важен поддял на биогеографията; Gaston et al., 2008/ по своя специален фокус на историчните причини, като допълнителни към селектиращите сили и другите екологични процеси, опериращи през повечето съвременни времеви рамки. Посредством своите иновативни генеалогични преспективи и своите значителни връзки с няколко различни области, филогеографията ще продължи да осигурява здрав емпиричен и концептуален мост между няколкото главни еволюционни дисциплини които имат ограничена комуникация помежду си. (Fig. 2).
Фигура 2. Генералното място на филогеографията и някой от нейните емпирични и концептуални свързващи функции, сред науките за биоразнообразието (modified from Avise, 2000).
Считана за една от най-големите войни на 20-ти век, този конфликт изправя Русия и Япония-доминиращите нации в североизточна Азия една срещу друга в битка за контрол върху Манджурия и Корея.
Но защо войната въобще избухва, и каква е историческата й значимост? Д-р Филип Тоул от университета Кеймбридж ще ви разкаже всичко което трябва да знаете за конфликта.
Кога и защо избухва руско-японската война?
Войната избухва на 8 февруари 1904. Руснаците и Японците се сблъскват няколко пъти преди това за контрол върху Корея и Манджурия, като и двете сили ги смятат за стратегически и икономически важни.
В началото на войната японския флот нанася неочакван удар по руския флот акостиран в пристанището извън Порт Артур, също така започва да спуска войски в Чемпуло, Корея.
Защо толкова малко от войната се помни във Европа?
Въздействието й е притъпено от по-големия конфликт който избухва във Европа 10 години по-късно. Но все още тази война се помни в Япония и Русия. Това, че не е известна не значи, че не е била важна в политическо или военно отношение.
Кой е спечелил войната?
Японците печелят всяка битка. Руските военноморски сили базирани в Порт Артур са разтърсени от загубите претърпени през Февруари 1904, и дори повече от смъртта на един от най-харизматичните им лидери, адмирал Степан Макаров, когато кораба му е взривен от мина през Април 1904.
Корабите им прекарват много от времето закотвени в пристанището преди да се опитат да избягат на 10-ти август 1904 около Корейския полуостров към Владивосток-най-източната руска тока. По време на разразилата се битка в Жълто море, флагманския кораб ‚Царевич‘ е понесъл тежки щети и бойната линия изпада в объркване преди да се оттегли към пристанището.
В крайна сметка флотата е унищожена от японските сили обсадили Порт Артур откъм сушата, и самия пристанищен град се предава през Януари 1905. Руснаците пращат още една флота към далечния изток която да балансира ситуацията, но и тя бива унищожена в битката при Цушима през Май 1905
Японската сухопътна кампания била ли е успешна?
Почти: Японците настъпват към Корея през реката Ялу в Манджурия и когато руснаците опитали да се бият, японците използвали фланкиращи маневри и ги принуждавали да отстъпват на север.
Двете най-важни битки са в района на Лиаоянг през Август 1904 и Мукден през Март 1905, и двете спечелени от японците чрез фланкиращи маневри, доброто качество на артилерията им и превъзходния морал и обучение на войниците.
Какво е историческото значение на войната и какви са дълготрайните й ефекети?
Японскатапобеда поощрява национализма в Азия и Африка. Японско-руската война е първата такава, в която не европейска нация побеждава велика европейска сила използвайки модерни технологии.
Парадоксалното е, че войната показва Японските имперски амбиции в Азия, по време когато европейските сили започват реално да оценяват националистките заплахи за техните империи. Японците успяват да установят контрол над Корея след войната, но срещат упорита партизанска съпротива от страна на корейците, което води до горчива омраза която и до днес трови японско-корейските отношения.
Войната издига и статута на САЩ в международните отношения. Президента Теодор Рузвелт събира конференцията в Портсмут САЩ която довежда войната към нейния край през Септември 1905 и печели на президента Нобелова награда за мир.
В Русия, войната води до бунтове които предсказват революцията от 1917, и показват колко политически дестабилизиращи могат да са загубите в бъдещи конфликти. Единствената част от руската империя която се е представила добре във войната е Транс Сибирската железница, която е доставяла припаси на стотици хиляди войници намиращи се далеч от Санкт Петербург.
Сблъсъка показва и знаците на положение на безизходица което ще се появи и по време на Първата световна война, но по време на руско-японската това положение не е чак толкова силно изразено тъй като армиите са по-малки и японците превъзхождали руснаците.
Войната показва и как се изменят законите на войната и хуманитаризма. През 1904-05 година японците третират руските пленници със респект, но по време на Втората световна война те не се чувстват обвързани със такива ‚западни‘ обичаи.
Руско-Японската война е едно ярко начало на 20-ти век, което показва как центровете на сила се изместват към САЩ и други не европейски държави, и колко крехки монархиите са станали.
Научната парадигма е обществен договор за устойчивост на понятията, чрез който учените от дадено поколение или няколко последователни поколения изследват явленията и процесите в даден клон на науката, като ги анализират и обобщават в полза на развитието на обществото. Т.е това е рамката, в която се осъществява главната цел на науката, която от създаването й от древни времена до днес е да бъде основен двигател на човешкото развитие и на човешкия прогрес. Според академик А. Акимов всяка научна парадигма се основава на първо място на развитието на физиката като основа за развитието на философската наука.
Общественият договор представлява физичната програма за развитие на всеки клон от науката. От рамка, в която учените изследват света, в един момент на насищане на знанията от този ъгъл на виждане, който е характерен за дадения етап на развитие на науката, научната парадигма се превръща в затвор, клетка, в която остарелите идеи търсят не само нови привърженици, за които са отворени всички врати, но и се отрича всяка новопоявила се мисъл за различно научно отношение или интерпретация на съществуващото дотогава. Появяват се т.нар. паранаучни трудове, които не се приемат, без даже да са прочетени. Утвърдените авторите в дадена област стават истинска спирачка на прогреса. Парадоксът е, че всеки изявен учен претендира за ново отношение, с което е дошъл като изследовател в лоното на науката. Но истински ново е онова откритие, което формира появата на нов научен мироглед. А то винаги е свързано с промяната във физичната основа, която разкрива тайните на света.
От една страна в това негативно отношение към различното има смисъл, защото обществото изисква задълбочено проникване в същността на процесите и явленията в конкретно избраната посока на развитие дотолкова, доколкото тя е важна за развитието му. А за това е нужен научен и времеви потенциал. Или, казано по друг начин, така се дава възможност за развитие на дадена идея и нейните конкретни планове и характеристики.
От друга страна обаче консервативното отношение в науката е опасно, защото при поява на революционна идея, тя очевидно, по силата на договора, ще бъде спряна. Според едно телевизионно изказване на акад. Акимов по БНТ (програма „Всяка неделя” с водещ К. Кеворкян) досега в научната практика новата идея се налага в чисто практически план не толкова в момента на появата й в обществото, колкото със смяната на научното поколение, което е утвърдило позициите на тази научна парадигма. Днес според него ситуацията е различна. Нуждата на човешката цивилизация, изправена пред глобална катастрофа, да промени научната парадигма е неотложна. Това налага нейната промяна преди смяната на поколенията.
За неустойчивостта на понятията. Условност при определяне на тяхната устойчивост въз основа на съществуващата физична парадигма. Що е неустойчивост? Променлива величина ли е тя?
Съществува връзка между промяната в понятията и семантиката на думите. Това взаимоотношение се наблюдава предимно когато се сменя научната парадигма. Ето следния пример: според информациологията дадено понятие се изследва в неговия план. А досега съществуващата гледна точка е изследване от даден ъгъл. Или от сбор от такива ъгли. Каква е разликата? Тук, разбира се, се вижда, че това са два подхода в усвояване на действителността.
Досега известният ъгъл на виждане на дадено понятие включва външното наблюдение, наблюдението отстрани. Самото понятие за обект променя срещата между т.нар. обект и т.нар субект – учения. Субектът – изследователят – се поставя в ролята на независим наблюдател. А обектът на изследване – на пасивния участник в комуникацията. Според теорията на информационното поле наблюдателят не е просто този, който отчита съществуващите дадености, но със самото си наблюдение спрямо обекта, той го променя (теоремата на Бел за нелокалната връзка. Както и наблюдението върху даден обект (вече условно наречен обект) променя и наблюдаващия го. Осъществява се информационен обмен на фино физично ниво. Оттук нататък, както обяснява теоремата на Бел, тези два обекта завинаги се променят. Възможността да се срешнат е осъществена. Друг е въпросът коя точно възможност е тя. Пак според информациологията възможностите за осъществяване на среща между два условно наречени обекта в пространството са неизброими, а се осъществява един в едно пространство. Въпросът е колко паралелни светове съществуват и как те си взаимодействат в общия еволюционен процес. Дали в този процес не действа т.нар. „ефект на стотната маймуна на Уилсън”, или това е все още непознат за нас закон? Така че ъгълът на виждане е достатъчно тясна рамка, за да се характеризира днес с него изследователският процес.
Кое е ново и кое е старо в науката и как да разбираме пътя на думите в науката.
Още древните гърци (9) са възприемали голяма част от основните положения на информациологията. Съществуването на общо поле, което съдържа информация за всичко преди и всичко след това са приемали и представителите на обективния идеализъм. „Представите за това световно информационно поле като «база данни» има дълбоки корени във философията на Платон, Лайбниц, Шелинг, Хегел и други представители на обективния идеализъм.”(9)
Може ли в такъв случай да се говори за нова теория, нови понятия, нова парадигма, когато и древните тайни (езотерични) науки изглежда, че дума по дума обясняват откритията на съвременната физика?! Или – защо науката да бъде информациология, а не – някои от предишните названия на онези научни феномени, които са известни от древността до днес? Когато човек има основно понятие от законите на информационното поле – и преди всичко основния закон за еволюцията, той неминуемо ще се съгласи с твърдението, че два пъти една и съща река не тече от едно място. За да се обясни създаването на новия клон от науката – информациологията – като съвременен феномен, трябва да се има предвид, че съзнанието на съвременния човек и съзнанието на нашите предци е различно. Т.е. базата на нашето разбиране за същността на информациологията е нашето израснало съзнание.
Успоредно със знанието и с много по-бързи темпове върви еволюцията на познанието – т.е. на всеобхватното знание. То съдържа в себе си знанието и опита на физичния ум, на емоционалния ум и на интелектуалния ум на човека. И като тяхна организираща и събирателна – духовното виждане у човека. Последното никога не е било самостоятелно или нещо извън еволюцията на невидимия свят. То се ръководи оттам. Световете на високите вибрации са достигали до нас с притежаваните от тях безценни знания за света и Земята по трансцендентален път, чрез който все повече хора умеят да го възприемат. По този начин са предадени основите на Нютоновата механика, както и квантовата физика и всяко друго базисно знание. В помощ на човека и усвояването на земния живот от него. Така са се появили и езотеричните (тайни, непознати) науки, също както и пророчествата на Нострадамус, които са всеизвестна, но не и необичайна практика за извличане на знанието от невидимия свят.
Целта на съществуването на човечеството на Земята, според някои учени, е била вероятно чрез усъвършенстване на неговия физически, емоционален и ментален свят да се постигне безсмъртието и на тялото му. Последните столетия от развитието на човечеството са белязани с развитието на менталния му ум за сметка на физическия и емоционалния. Древният човек е възприемал света линейно. Неговите представи са стигали дотам да регистрира даденостите като линия. По-късно се формира разбирането му за повърхност, площ, място. Той възприема света и Земята като плоскост с дължина и ширина. Тези представи са напълно в синхрон със знанията, заложени в геометрията на Евклид, на чиято база Нютон създава своята механика. ”Тук етерът е някаква еластична материална среда, която осигурява взаимодействието на телата, които притежават гравитационно поле. Съгласно теорията на Нютон само благодарение на наличието на етера се осъществява притеглянето на телата чрез силата на гравитацията”.(1) Човекът, излизащ от Средновековието се пробужда към триизмерността: дължина-ширина-височина. Нещата вече имат своя обем. Пространството изглежда преодоляно от нашето съзнание. Но днешният човек знае, че има и четвърто измерение* – на времето, в което предметите, обектите не са едни и същи в два различни момента, т.е. две различни времеви точки. „Тези представи са претърпели кардинални промени и по трите позиции, когато в началото на ХХ век беше формулирана теорията на относителността на Айнщайн… геометрията на Евклид беше заменена с геометрията на Риман; – понятието етер беше заменено с понятието физически вакуум.”(1)
В този порядък трябва да се разглеждат от съвременния учен и древните, и съвременните достижения на човешката мисъл. Те не са фиксирани като едно единствено познание, защото знанието е резултатът от възприемането на действителността чрез нашето съзнание (взаимодействието ние:светът и светът:ние). А не някаква „обективна истина”, понятие, което лесно може да бъде оборено. „Досега материалистичният подход винаги претендираше за научност, т.е. обективност, каквато не съществува. И със силата на предварително договорения конкретен принцип за научност – научната парадигма – ставаше недосегаем. В резултат този подход представлява налагане на определен (частен – от гледна точка на взаимосвързаните системи) историко-културен или прагматичен поглед върху известното събитие. Чрез него се налагаше не само в науката, но и в публичното пространство т. нар. обективна оценка на събитието. Тя се обяснява като наблюдение върху обекта на научното изследване, което не засяга и не се влияе от субекта, провеждащ изследването. Оттам и името на метода – обективен, неповлиян от изследователя. Но квантовата механика доказа, че на квантово ниво резултатът еднозначно зависи от изследователя, т.е. от човека, който го изследва. Следователно, да се натоварва с обективност даден извод, без да се отчита кой го е направил, просто е невъзможно.”(3)
Както се разбра по-горе, съзнанието ни през всички тези еволюционни скокове е било в различен стадий на развитие. В този план се разбира и един друг аспект от информациологията – как въздейства или още повече взаимодейства нашето съзнание със съществуващите понятия и обекти от действителността. Теоремата на Бел ни отвежда в една фантастична реалност на връзка завинаги между два обекта, които са се срещнали само веднъж. Тя обяснява паметта на информационното поле и теорията за фантомите, които остават след думите и делата на човека и всичко в невидимото пространство или по-точно – в невидимия свят. Светът на по-високите вибрации.
Едно познание освен това не може да бъде едно и също в два различни еволюционни момента. Еволюцията е всеобщ закон на Битието. И ако едно знание се е възприемало по един начин само преди сто години, днес вече нещата са други не само поради факта на нашето пораснало съзнание, а и поради развитието на всички останали обекти от Вселената. Пример в това отношение може да бъде понятието „енергия”, което все още се възприема от адептите на Бялото братство като единственото обяснение на развитието на света. Но науката обяснява, че онова, което се движи със скорост милиарди пъти по-бързо от светлината, не може да бъде енергия, защото не притежава маса. Става въпрос за информация. И информационно поле. И, разбира се, за основите на информациологията.
Понятието „информация”. Значението на това понятие изключително много се е променило през вековете. Например в електронната библиотека Уикипедия четем: „Информация (от лат.Informare – обучавам) първоначално е означавал процесът на предаване на знание или съдържание. В днешно време под информация се разбира налично, използваемо знание. Информацията е сведение или данни за каквото и да било събитие или обект”
Първоначално се е определяло като сведения, данни, представи. (12), а днес според специализирания речник по информатика (12) от 1975 г., издаден в Москва на 13 езика, има повече от 160 понятия, произлезли от първообраза на латинската дума – “information”. А Ж. Винарова и М. Вуков определят класическотото схващане за информацията досега като: „сведение или съобщение за нещо или за някого, предназначено за осведомявани или осведомяващи се.” Интересно и пълно изглежда и тяхното определение, което те дават в учебника си „Информационни системи в медицината и образованието”: Информацията е сведение за обекти, процеси или явления от околната среда, за техните параметри, свойства и състояния, които се възприемат от двете категории информационни системи – живи организми или управлявани машини, в процеса на тяхната жизнена дейност или работа. Но в момента информационната физика приема, че според науката за информационното поле всеки обект от Вселената, наричан преди жив или нежив, притежава полето на душата, т.е. собствено вторично информационно поле. Това значи, че понятието за информация не различава тези две категории като две, а като един информационен обект. Дори само този факт говори, че еволюцията на това понятие в продължение на не повече от 50 години е вървяла с космически темпове. От понятие, което означава данни, събиране и интерпретация на данни, процес на взаимодействие – отразяване на действителността през погледа (познанията) на възприемащия информацията – в понятие от физиката, което означава поле, фино материално поле, и обект, който излъчва фино материално поле, без значение дали е биологично жив или – не.
Пак същите автори дават извлечени петнайсет определения от уебпространството, които общо взето повтарят разбирането за информацията като събиране, набор и интерпретация на сведения, данни, факти от ползващия ги.(4)
Едва в средата на миналия век – през 1948 г. американецът Клод Е. Шенън (12) говори за количество информация и теория на информацията. Момент, в който вратите на науката се отварят широко за използване на понятието като научен термин.
Бащата на кибернетиката Норберт Винер дава определение (12), което звучи повече като философия, отколкото като определение, дадено от математик: „Означение на съдържанието (на сигналите), получени от външния свят в процеса на приспособяването ни към него и приспособяването на нашите чувства.” Ако сравним определението на акад И. Юзвишин*(„Информация – това е генерализационен безначално-безкраен единен законопроцес на микро- и макромерните отношения, взаимовръзки и взаимозапазване на енергията, движението и масата на основата на резонансно-клетъчната, честотно-квантовата и вълновата природа на светлината, топлината, звука и другите свойства и форми в микро- и макроструктурите на Вселената.” И. Юзвишин „Информациология”), ще разберем, че това определение е най-близо до определението на торсионната физика за информация от всички по-горе дадени или споменати определения. Защото има пренос на съдържание, както и че ползва нас, хората. Само че светът, предложен от Винер като външен (респективно – и вътрешен – нашите чувства и интелект, които се приспособяват към съдържанието) практически е едно единно цяло – първично торсионно поле, в което се съдържат плановете на всички обекти във Вселената. То се задвижва от мисъл или чувство, т.е. от движението на наблюдателя. И не се отнася само за хората, а за всички обекти във Вселената. Норберт Винер твърди, че: „Информацията е информация, а не – материя или енергия.” Според Е. Акимов (1) – за „особената форма на материята, наречена физически вакуум” – „Тази среда е материална, но не веществена. Това е полева, информационна структура.” И за нивото на полевата материя – „…взаимодействието е такова, че те (торсионните вихри –бел. моя) пренасят не енергия, а информация.” Когато става въпрос, че информацията е „форма на отражение” (4,12) , „процес на взаимодействие”, която няма материална същност”, абстракция, информационната физика приема, че това е особена форма на движение на най-фината материя, която поради ниските минусови материални стойности не се зачиташе досега от науката като практически използваема. Но информацията е наистина взаимодействие, предизвикано движение на информационни кластери между структурните обекти в едно ниво или между различни структурни нива на информационната йерархия във Вселената. Хр. Туджаров (13) я описва като първична и вторична: „ Първична информация – информация получена от една или няколко системи, която се обработва от друга система, с цел получаване на нова информация. … Вторична информация – всичко, което се получава в резултат на обработка на първична информация и данни.” Според информационната физика има два вида полета на информацията – първично информационно поле – общото поле на съзнанието на Вселената и вторично торсионно (информационно) поле – информационното поле на всеки отделен обект във Вселената, който е част от дадено структурно информационно ниво.
Ж. Винарова и М. Вуков (4) се насочват актуално към разглеждането на информацията като „извънтелесна” и „вътретелесна”. Вече стана въпрос по-горе, че информацията не може да бъде външна или вътрешна. Тя е вездесъща. Прониква навсякъде и във всичко. От всяка клетка, система, процес и видимите тела на човека до неговите невидими параметри. Но от друга страна приемането на „вътретелесната” информация като обект за разглеждане от медицинската наука е особено важно за човек днес. В това отношение има събрани достатъчно изследвания, фактически материал и доказателства за съществуването на човека като информационен обект – копие на матрицата на Вселената.
Във философията понятието за информация вече е основа за разбиране на устройството на света от гледна точка на фините (минусовите –от порядъка на 10-¹²) (15) материални полета. За да се разбере основната разлика в използването на понятието, ще съпоставим някои определения, използвани от учени, които оставят следа в разбирането за това понятие.
Според Д. Томов (12) в труда си Въведение в научната информация и информационната култура класифицира информацията според сферата на нейното възникване като: „елементарна, възникваща в неживата природа; биологическа, възникваща в растителния и животинския свят с разновидността й генетическа информация и социална, възникваща в човешкото общество.” Според Г. Шипов (15) информацията е пренасяното от полевите вихри на торсионните полета, чието категорично място е финият материален свят – светът на идеите и плановете за всички по-груби материални нива: на елементарните частици, газовете, течностите и твърдите тела. В човека се съдържат всички основни нива на йерархията на Вселената – твърдо тяло, течности, газове, плазма, елементарни частици, фини астрално, ментално и казуално тела. Съдържа се и най-високото ниво – седмото, чрез което информацията на Божественото начало достига до нас. Информацията може да бъде в статично състояние или да прави пренос в зависимост от факта дали някой предизвиква физическия вакуум. В този смисъл теорията за отражението на информацията от гледна точка на диалектическия материализъм (12) е само частен случай на нейното движение. Още повече разпространението на информацията не зависи от други фактори и не е нито еднопосочно (отражение), нито двупосочно |човек:информация или информация:човек|. Разпространението на информацията е многопосочно или още по-точно вездесъщо по причина на факта, че тя се разпространява до всяка точка на Вселената в мига на нейното възникване. Или според холографското свойство на Вселената всеки обект съдържа в себе си информацията за цялата Вселена във всеки един момент. Съответно и човек има достъп до цялата информация, която се съдържа във Вселената. Протичането на информационния обмен става със или без съгласието на човека чрез неговите фини информационни тела.
Промяната в трактуването на понятието за информация от външен, нематериален и необвързан със същността на физическите закони обект на човешкото знание във вътрешен, част от човека и Вселената главен компонент на всичко и всички, който наистина не притежава енергия, т.е. не е веществен, но представлява фино материално поле, което се подчинява на определени закони, придобива все повече известност и значимост след включването му от ООН в регистъра на науките през 2000 г. Това практически означава и официална промяна в научната парадигма, която, парадоксално, вече е утвърдена, но не е позната на много университети в света.
Следователно познанието вече не означава натрупаните книги и битове информация в различен видим за човешкото око материален и веществен вид, а конкретното постижение на еволюцията (нейният невидим свят и видимото му проявление) в един конкретен момент. Както и вечно съществуващото ниво на плановете, идеите за следващи и предишни моменти, т.е. всичко заложено в първичното информационно поле.
Познанието въздейства всеобхватно (видимо и невидимо) върху хората и другите обекти във Вселената по един и същи начин – с цялото си енергоинформационно поле. Те се променят в зависимост от конкретните планове на еволюционния процес за всеки от тях и в рамките и на правото на избор за действие и съответно допринасят за създаването на информацията – процеса на еволюцията. Полето, което управлява еволюцията, е полето на всезнанието, познанието и съзнанието на човечеството. А хората като общо съзнание (5) винаги „управляват” това познание, въздействайки му от своя страна с нивото на своето съзнание. Т.е. еволюционният процес пак има потенциал на възможните варианти в зависимост от плановете на Твореца.
Например онова, което е дадено в Библията, днес се възприема по друг начин благодарение на факта, че сме достигнали определеното ниво за определеното познание. Така за хората днес е възможно да надникнат по-надълбоко и да се опитат да разшифроват (според нивото на съзнанието си) Библейския код, нещо невъзможно в такава степен в миналото, защото духовността на съвременния човек е много по-развита, по-висока от тази на неговия предшественик. В този ред на мисли информацията за еволюцията на информационните центрове на човека, т.нар. чакри (фокуси на енергоинформационните тела на човека) доказва тази теза. (8) Докато първобитният човек е имал развита първа (отговаряща за оцеляването) и втора чакра (отговаряща за възпроизводството), днешният човек е достигнал до развитието на пета чакра, отговаряща за творческото развитие, а някои хора развиват и шестата – на висшата духовност.
Условността в установената устойчивост на понятието за информация днес като основа за развитието на новата научна парадигма има смисъл дотогава, докато изясни, чрез физиката и всяка друга наука, резонансните аспекти при взаимодействието между човешкото съзнание и познанието за информационното поле.
Връзката между промяната в понятията и семантиката на думите е заложена в самия еволюционен процес. Първо се променят идеите (понятията), които могат да се установят в даден етап с определени думи. Те пък стават носители на смисъла, на идеята и конкретното съдържание. Понякога и често пъти науката стига до определени характеристики (страни) от съдържанието на едно понятие. В случая – понятието информация. И то получава своето име – дума.
Второ, името се дава, запълнено само с част от съдържанието. В случая с думата информация съдържанието й до появата на теорията за информационното поле (15) е едно, а с теорията се стига до нова идея, която не просто допълва съдържанието на дотогавашното понятие информация, а „отваря” всички прозорци и врати за науката за нов прочит на Битието. В който процес главната въпросителна е резонансът на човешкото съзнание със света, който твори и сътворява, взаимодействайки с Вселената. (11)
Успоредно с развитието на човека и неговото общество се развива и науката, която изконно е призвана да помага на човека в усвояването на познанието. Това е и причината новата стара наука за света да се нарича информациология, наука за информационното поле, а не „обективен идеализъм” или друго название от терминологията на езотериката. Човек е достигнал до познанието точно на информационното поле и то не само обяснява древните познания на хората, но ги разкрива толкова пълно, колкото е нивото на развитието на човешката мисъл и духовност, т.е. на човешкото съзнание днес.
Остава въпросът как учените да използват занапред това понятие при неговата пораснала значимост, при драстичната промяна в неговата знакова система, при богатото разнообразие от значения, някои от които вече влизат в смислово противоречие – даже само от даденото като примери дотук. Очевидно е, че предстои по-задълбочено и всобхватно систематизиране, което да даде правилните насоки в използването на понятието информация в полза на еволюцията на човешкия род.
В заключение може да се твърди, че промяната на парадигмата днес е в пряка връзка с промяната на съзнанието на хората, а оттам и промяната на „познатите” непознати понятия и техните имена.
Литература:
1. Акимов 1999: Акимов Е. Облик физики и технологий в начале ХХІ века. Выступление на научно-педагогической конференции ”Идеи Живой Этики и Тайной Доктрины в современной науке и практической педагогике” в Экатеринбурге, Москва, изд. Шарк ;
2. Акимов 1996: Акимов Е. – Физика признает Сверхразум – сп. Чудеса и приключения.
3. Александрова 2007: Александрова Р. Невидимите хоризонти на славянския корен или – преди да обикнеш другия, научи се да обичаш себе си Сб. „Славяните и Европа”, съставители М. Младенова и Е. Дараданова, София , изд. „Херон прес”
4. Винарова, Вуков 2005: Винарова Ж., Вуков М .Информационни системи в медицината и здравеопазването, С., изд. „Сиела – софт енд паблишинг”
5. Грабовой 2003а: Грабовой, Гр. Прикладные структуры создающей области информации учебное пособие. М., ”А. В. Калашников”, 2003.
Грабовой 2003б: Грабовой, Гр. Унифицированная система знаний. М., ”А. В. Калашников”, 2003
Грабовой 2001: Грабовой, Гр. Ирациональные методы предотвращения глобальных катастрофических процессов, представляющих угрозу миру. [Лекционен курс.] М., 2001.
8. Пучко 2003: Пучко, Л. Г. Биолокация – приложна радиестезия. С., 2003
Войната в офиса ви е позната, нали? Онази тихата, ползваща всички средства – клюки, междуособици, заплати, квалификация, кадърност…
Как да останем в периферията на горещото ядро от неважните за работния процес неща? Как да съсредоточим екипа си върху работните проблеми, ако сме начело на някакъв отдел – без значение какъв – когато работата с хора е най-трудното нещо на този свят?
Безброй са изследванията и публикациите за отношенията в офиса, за екипната работа, за „добрия лидер”. Но колко от мениджърите всъщностса Лидери? Колко от хората имат време да се занимават със сухи извадки и предъвкани факти? И най-важният въпрос – колко души им вярват?
Ще обърнем внимание на самата реалност – как всъщност се развиват взаимоотношенията в един екип между ръководител и подчинени. Целта е да се представят нещата такива, каквито са и простичко да се обясни какво всъщност разделя или събира даден екип и как можете да стигнете до главното “Л”.
За да управлява успешно и ефективно един мениджър, той следва да осъществява основните функции, които са присъщи на мениджмънта, а именно: да планира, да организира, да координира и т.н. Това са функциите определени от, най-общо казано, неговата длъжностна характеристика. Да обърнем внимание, обаче, на едно друго качество – способността му да комуникира с подчинените си. Това си е цяло изкуство и всъщност от неговото усвояване зависи доброто сработване на екипа и в крайна сметка – положителните резултати на целия отдел.
Професор Фред Фидлър* е убеден, че е по-лесно да се промени всичко в работната ситуация, отколкото да се промени личността на човека и оттам – неговият лидерски стил. Той вижда ефективността като промяна на ситуацията, а не като промяна на стила. Да се промени стилът на ръководителя чрез обучение според Фидлър е изключително трудна задача. Бих допълнил, че докато функционното ръководство може да се изучи и приложи в някаква стереотипна схема, качеството „комуникация” е по-скоро вродено умение. Манипулирането на моментните ситуации е доста полезно, за този който умее да жонглира с наблюдението, анализа и практическите приложения на изводите. Ако имате проблем с управлението на екипа си, може би трябва да обърнете менталния си взор към осъзнаване на следните действия, които трябва да предприемете:
даване на ръководен пример
защита интересите на подчинените
интерес към задачите на всеки подчинен
помощ при изоставане от графика
наблюдение на взаимоотношенията между служителите
умение за запазване на „шефски” авторитет
Казвам „осъзнаване”, защото всеки е склонен да си интерпретира тези понятия по свой начин, без да се замисля какво значат те за хората, с които работи. За да синхронизираме нещата, ще ги разгледаме едно по едно.
Ръководният пример.
Трябва да знаете едно нещо – дори и преди да сте били част от екипа, на нивото на останалите, хората ви вече гледат на вас с друго око. Те виждат във ваше лице човек, който взема повече пари. Човек, за когото по презумпция смятат, че работи по-малко от тях. Това, разбира се, кара мнозинството да не си дава “много зор” по задачите или най-малкото – да ги върши до минимум необходимото. Как ще накарате тези хора (в един екип те винаги са налице) да се мотивират и да дадат максимума от себе си? „Доброто” старо крещене и мъмрене не помага, ако си имате работа с разумни хора. Заплахите за уволнение при най-малкото провинение са стереотип на една минала епоха. Вие си комуникирате с човеци, те разбират и от други неща, освен от прекомерната директорска показност. Не се опитвайте да ги смачкате с властта си (още повече, че тя все пак не е толкова голяма), защото това ще е краят на вашето водачество.
Сутрин винаги идвайте пръв на работа, а вечер по възможност си тръгвайте след подчинените си. Никога не го натяквайте на останалите, те просто винаги ще се чувстват леко гузни за това и ще спечелите уважението им като точен и ефективен тип шеф. Не толерирайте обаче закъсненията. Може да не обръщате внимание, когато няма спешна работа, но при гонене на срокове, трябва всички да са наясно, че това влияе негативно на целия екип. Можете да го обясните на хората не на оперативка, а примерно на групов обяд. Там те биха приели думите ви не като студена заповед, а по-скоро като сключване на пакт, който ще спазват.
Нека всичко това не ви звучи като глупав план за подмазване на подчинените си. Това са функциите на лидера – да покаже, че може, желае и прави повече от останалите, защото хората не се нуждаят толкова от някакъв директор, отколкото от водач. Както отбелязва Питър Дракър*, единственото определение за лидер е “някой да има последователи”. Още от племенната система в света е валидно следното правило – всички следват човек, когото харесват, когото признават за нещо повече от себе си, а не такъв, когото презират. Човек е водач, когато другите го признаят за такъв, а не сам да е надянал короната. Това е най-естествената мотивация за служителите ви – да приемат, че са в един състезаващ се отбор, а вие сте негов капитан.
Протекция на служителите.
Мотивацията трябва да се захранва и от още нещо, което е задължително, за да бъдете добър водач за хората си – защитата на интересите им. Знаете как от страна на други отдели се появяват нездрави опити да се хвърля сянка на съмнение върху способността на отделни членове на екипа или върху качеството на работа. Това е двоен удар – веднъж за екипа, втори път за вас самите. Длъжни сте да защитите имиджа на отдела, като отхвърлите тези съмнения пред водещите мениджъри на организацията. Това може да направите по няколко начина като:
споделяте с шефовете колко добре се сработвате с екипа;
хвалите отделни членове на отдела;
ги държите постоянно в течение на текущата ви работа.
Така „зломислителите” ще бъдат разбити с истината, без да се налага да сплитате и вие интриги.
Друг момент в протекцията на подчинените ви е отстояването на техните работни права. Щом вие сте позволили отпуск, например, не може друг, пък било и по-висш мениджър, да го отменя. Това уронва престижа ви и кара служителите ви да се съмняват във вашата дума. Дайте на останалите мениджъри ясен знак, че отделът е ваш и не търпите вмешателство. Заплати не можете да коригирате, но е добре, когато някой се справя перфектно и с желание се хвърля в изпълнение на задачите, да го предложите за минимално увеличение на заплатата или поне за еднократен стимулиращ бонус.
Понякога наистина някой бърка и тогава вие можете да бъдете смъмрен за пропуските на екипа. Не хвърляйте вината веднага върху колегата си, защото: първо това не е спасителен пояс за вас, второ – жалко е. Всеки бърка, дори и вие, така че поговорете с този ваш подчинен и той ще си вземе бележка. Не забравяйте, че много по-лесно е за шефовете да уволнят не вас, а някой от екипа ви. Не позволявайте да ви се понесе славата на ръководител, който с лека ръка оставя служителите си на улицата. Естествено, ако той продължи с тези „изцепки”, то време е да се разделите с него, тъй като или го прави нарочно, за да ви навреди, или е некомпетентен. И в двата случая не е нужен на екипа.
Интересът към персоналните задачи.
Никой не харесва шефа, който се „овесва” на врата на подчинените си и ги следи като гестаповец докъде са стигнали с работата си. И напълно резонно – работоспособността на човек по-скоро ще спадне, отколкото да достигне някакви положителни стойности. Никога не изнервяйте по този начин екипа. Хората не обичат да бъдат дебнати. Не харесват, когато някой застава до тях и половин час се взира в монитора им, примерно. Все пак, това са хора, които се имат за специалисти – ако се не се съмнявате в тях, защо ги следите? Това се питат те и определено смятат, че си е чиста проба тормоз.
Има по-добър начин да сте в крак с постигнатото от екипа. Похвалете всички заедно, ако работата върви, покажете им, че тя е важна за организацията, в която работите, накарайте ги да се отпуснат. Никой няма да ги разпъва на кръст, те ще се държат естествено. Така в самия процес на работа, подчинените ви сами ще обсъждат с вас и останалите си колеги тяхната част от проекта, етапите, по които работят, без да имат чувството, че са разпитвани. Изхождайки от максимата, че в спора се ражда истината, ще добавя, че с разговори се постигат по-добри резултати. Така ще имате време да изгладите някои пропуски, които иначе – в последния момент, ако не знаете за тях – ще си останат като минус за отдела.
Когато поставяте задачите на отделните служители, постарайте се това да не прозвучи като фараонска заповед. Не забравяйте, че всички сте там на работа; никой не е роб, за да се държите надменно с него.
Помощта при нужда.
Не всички могат да се мобилизират винаги на сто процента. Има фактори, влияещи негативно на всеки, дори и да е добър служител по принцип. Ако някой започне в даден момент „да дава фира”, вие сте длъжни да му помогнете. Поговорете с него насаме. Не подхващайте темата с това, че ще му “ударите рамо”; по-добре му кажете, че имате свободно време и искате да вземете част от работата му, за да не бездействате. Хората обикновено имат самочувствие, а така вие няма да го засегнете. Ако човекът гледа обективно на нещата, ще приеме с удоволствие и ще оцени помощта, още повече, че останалите от екипа няма да разберат за моментната му слабост. Ако пък откаже, ще е длъжен да свърши навреме и сам, евентуално работейки извънредно.
Да обърнем внимание върху друг факт – онези, които системно не се справят със задълженията си и се налага постоянно някой друг да върши работата им. Такъв човек трябва да разбере, че всичко си има граници. Не трябва всеки път да карате останалите да му помагат, защото те не са виновни за неговата очевидна неспособност. Колкото и да е неприятно, тези служители трябва да бъдат предложени за уволнение. Все пак, това е пазарна икономика и не може вечно да се прикрива „дупката” на екипа. Понякога са нужни сериозни мерки, спасяващи работата, а не един нескопосан индивид.
Наблюдаване на служителите.
Не, не монтирайте скрити камери и микрофони, няма да се занимавате с изнудване. Вие трябва да наблюдавате служителите си, не за да ги хокате и овиквате в случай на грешки, а точно за предодвратяване на тези грешки. Какво искам да кажа?
Както и в училище (спомняте си го, нали!), така и в офиса има някакви вътрешни междуособици – тази била дебела, онзи пък простак, на трети жена му му изневерява… Да виждате нещо свързано с работата? Не, но такива са хората; много изследвания показват, че в офиса повече време се отделя за клюки, отколкото за съществената работа. Положението се влошава в момента, в който всичко това започва да вреди на екипа и да го дърпа назад. Този не иска да работи по проект с онзи, защото знае, че не го харесва, другият пък не се погажда с някого, по чисто политически убеждения… и още подобни проблеми. Натоварването предизвиква едно дребно раздразнение у човек, което впоследствие се натрупва и тези с по-лабилна психика гледат да си го изкарат върху околните. Работа, разбира се, никой няма да спре, защото е длъжен да я върши, но ефективността спада. Не е важно нещо да се направи, ако ще е през пръсти, важно е да бъде както трябва. Ето защо вие сте човекът, който щом разпределя задачите на екипа, трябва да вземе под внимание тези наглед дребни междуособици. Една от тях е смешна, три вече са пагубни за работата.
Никога не поставяйте обща задача на хора, които изпитват някаква неприязън един към друг, защото в процеса на работа, тя ще се превърне в омраза, което ще насади тягостна атмосфера в екипа. При възможност дори слагайте тези хора в различни стаи. Така всички ще са доволни, а най-главното – работата ще си върви нормално. Ако се появи „неизлечим” конфликт между някои от служителите, време е да си поговорите с тях като шеф с подчинени и да ги накарате да повярват, че е по-добре да си мълчат и да не изнервят останалите си колеги, ако не искат да си търсят друга работа. Това е доста краен вариант, носитуацията го налага.
Лидерският авторитет.
Ако нещо от дотук казаното ви е навело на мисълта, че трябва да угаждате всячески на всички, веднага ще ви поправя. Пагубно ще е за вас да тръгне слуха, че сте мекушав, още по-лошо – безконтролен спрямо подчинените си. Не забравяйте, че и над вас има началници, а вашата цел не е да ви заменят с друг.
Наполеон Бонапарт е казал, че стремежът да се властва над себеподобните е една от най-силните човешки страсти. Това е в природата ни… но как точно ще го правим е лично наш избор. И не е само въпрос на избор, а и на умения. Далеч по-добре е да наметнем кадифеното покривало на разумното ръководство върху желязното си желание за властна дисциплина.
Не позволявайте на служителите да си мислят, че те определят върху какво ще работят, кога ще ползват отпуск или с колко ще закъсняват за работа. При първо провинение на някого, говорете с него насаме, но при рецидив, постарайте се всички да разберат, че са работили повече заради него. Никога не викайте, това не са деца, но по самия ви тон хората трябва да знаят кой е шефа. Когато давате задачи не казвайте „ще свършиш ли това?”, предпочетете ясното „свърши това!”. В никакъв случай не позволявайте да се промъкне съмнение в ръководните ви умения. Трябва да осъзнаете, че тези хора са ваши подчинени, а не тайфа приятели от училище, и да ги накарате и те да го осъзнаят. Може преди да сте били колеги, но правилното функциониране на отдела не търпи прекаленото фамилиарничене. Не позволявайте разни хора да се позовават на познанството си с вас и да не си обслужват задълженията. Всички трябва да са наясно, че ако работата върви напред, можете да бъдете разбран човек, но ако нещата се объркват, ще сте строгия шеф. Всичко опира до чувството за справедливост на хората и осъзнаването на факта, че когато допускат грешки, сами са си виновни за наказанието, а когато се справят добре, ще бъдат поощрявани. Е, ако работите с идиоти, замислете се дали да не смените организацията.
Разбира се, има още много фактори, с които трябва да се съобразявате, но те произлизат винаги от описаните шест.
Трябва да се знае, че това не са задължителни правила, които да спазвате фанатично. Не твърдя, че важат за всички ситуации, но съм убеден, че зачитането им би имало позитивен харарактер върху изпълнението на ръководни задачи. Ако всичко това ви се вижда безсмислено и прекалено, или пък повърхностно и ненаучно – няма нищо страшно. Отделът и работата няма да изчезнат… И дали този перфекционизъм всъщност е важен?
Но ако сте се замислили след прочетеното, ще ви кажа кое е важно за мен: както преди сте положили усилия да се изкачите до ръководна функция, самоусъвършенствали сте се постоянно с голяма доза упоритост, така и сега да докажете и на вас самите, че ставате за лидер, заслужавате уважение и знаете как да постигат своите и тези на компанията цели. Вярата в себе си е изключително силно оръжие във всяко едно отношение. Трудно е за осъзнаване, но след това е лесно за използване в изкачването на вашата стълба.
Тогава ще дойде и главното „Л”.
*Фред Фидлър е пенсиониран професор по психология от университета във Вашингтон(модел на “непредвидените ситуации”)
”Бъдещето на фехтовалното изкуство е в неговото минало”
Автор: Здравко Пеев
През последните 15 години се наблюдава засилен интерес в световен мащаб към алтернативното изучаване на европейското средновековно културно наследство и популяризирането му по нестандартни начини.
Един от клоновете на това културно наследство е традиционното военно изкуство практикувано от средновековния човек или така наречената историческа фехтовка.
По какво тя се различава от това което можем да видим в една историческа възстановка или от спортната фехтовка?
Историческата възстановка е преди всичко занимателен спектакъл пресъздаващ конкретно историческо събитие а „боевете” в него се водят по строго определени, безопасни и опростени правила. Също така „оръжията”, които се използват са с различни характеристики и качества от историческите образци поради безопастността на участниците.
Спортната фехтовката е динамична атлетична игра в която при установени правила двубоят се печели с отчитане по точкова система. Инструментите с които се практикува не могат да бъдат наречени оръжия в исторически аспект.
В традиционната фехтовка правилата се свеждат до това, да бъдеш оцелелият след сблъсък с въоръжен противник. Иначе казано – приложно бойно изкуство. И няма как да е друго, след като целта на средновековните и ренесансови фехтовални учители е била да научат човек да се бори за живота си в различни случаи – на дуел, в битка, както и в битови ситуации.
Целта на организациите – клубове, школи и т.н. по историческа фехтовка, разпространени в Европа и Америка е в тях да се изучава традиционното средновековно военно изкуство за самозащита. А за да сме още по-точни – кодифицирана и методична бойна система за работа с исторически оръжия – различни типове мечове, кинжал, палка, копие и други полеви оръжия, както и ръкопашен бой. Информацията за тази бойна система е достигнала до нас чрез голямо количество запазени исторически документи – трактати по фехтовка от периода XIII-XVII век.
Именно тези подробно написани, а много от тях и илюстрирани книги са завета на фехтовалните майстори от миналото и ключът към възстановяването на това специфично и значимо културно наследство. Тези документи са разнообразни по произход – основно германски, италиански, английски, испански и др. Важно е да се отбележи, че в някои от тях е засвидетелствана пряка приемственост на традицията между поколения фехтовачи – майстор след майстор…
И така какво можем да видим, ако за пръв път отидем на тренировка по средновековна фехтовка в Школа „МОТУС”?
Със сигурност няма да намерите „рицари” в средновековни костюми, които да си разменят галантности. Ще видите хора, които отработват основните стъпки – един от най-важните елементи във всяко бойно изкуство. Други ще упражняват по двойки атаки сечейки и мушкайки – в главата, в ръцете, в тялото… Когато базовите атаки се усвоят добре, се преминава към контраатаки. По-напредналите работят в свободен спаринг с дървени оръжия, а други – с метални. Металните оръжия са затъпени. Но това е единствената разлика с техните исторически аналози. Теглото, размерите, динамичните качества на тренировъчният инструмент трябва да са еквивалентни на историческите образци за да не се изкривяват действията. Любопитна подробност е, че за разлика от популярното мнение, мечовете не тежат по 5 или повече килограма, а теглото им варира между 1000-1300 грама за едноръчен меч и 1400-2300 грама за двуръчен меч. Тренировъчните мечовете, използвани в Школа „Мотус” са прецизни реплики на средновековните образци.
Видът на трениращите включва още униформено спортно тренировъчно облекло и разнообразна екипировка – здрави ръкавици, фехтовални маски, налакътници, наколенници и др.
Основите на Школа за средновековна фехтовка „МОТУС” са поставени през 2003г. Тогава е създадена експериментална „Група за изследване и изучаване на средновековни бойни изкуства“. През 2006г. групата става фехтовален клуб и приема името „Мотус”, а през 2010г. се трансформира в школа.
В Школата се изучават три основни дисциплини – едноръчен меч, едноръчен меч и малък щит (бъклър) и двуръчен меч. Работи се също с копие, палка, тояга и други.
Дейността на „МОТУС” , освен стандартните 5 тренировки седмично, включва демонстрации, семинари, участия и консултация за филмови продукции и други.
Най-важното събитие в годишният календар на Школата са традиционните летни тренировъчни лагери, където в рамките на десетина дни, в приятна атмосфера, се тренира особено интензивно. Лагерите са отворени и за всички, които имат интерес към средновековните бойни изкуства, но не са трениращи в Школата.
Едно от най-емблематичните места, където Школа за средновековна фехтовка „Мотус” е „завоювала позиции”, е Националния Военноисторически Музей, където се провеждат ежегодно традиционни демонстрации по случай Георгьовден – деня на храбростта. Националния Исторически Музей също е бил домакин на демонстрация на Школата.
На 12.02.2011г. Школа „Мотус” взе участие в едно безспорно уникално за нашата страна събитие – състоя се демонстрация по покана на „Фехтовална Академия Васил Етрополски”, където под един покрив публиката можеше да види разликите между спортната и традиционната фехтовка.
Информация за тренировките и всичко свързано с школата може да откриете на нашата фейсбук страница – http://www.facebook.com/MotusSwordsmanship
Както и да видите наши видеоматериали в YouTube – http://www.youtube.com/user/MotusSwordsmanship
За контакт: zdr_peev@abv.bg; motus_freifechter@abv.bg
Този сайт използва ‘бисквитки’ (cookies), за да ви предостави възможно най-добро потребителско изживяване. Можете да промените настройките си за бисквитки, или в противен случай приемаме, че сте съгласни с нашите условия за ползване.ПриемамПрочети повече
Правила на поверителност
Privacy Overview
This website uses cookies to improve your experience while you navigate through the website. Out of these cookies, the cookies that are categorized as necessary are stored on your browser as they are essential for the working of basic functionalities of the website. We also use third-party cookies that help us analyze and understand how you use this website. These cookies will be stored in your browser only with your consent. You also have the option to opt-out of these cookies. But opting out of some of these cookies may have an effect on your browsing experience.
Necessary cookies are absolutely essential for the website to function properly. This category only includes cookies that ensures basic functionalities and security features of the website. These cookies do not store any personal information.
Any cookies that may not be particularly necessary for the website to function and is used specifically to collect user personal data via analytics, ads, other embedded contents are termed as non-necessary cookies. It is mandatory to procure user consent prior to running these cookies on your website.









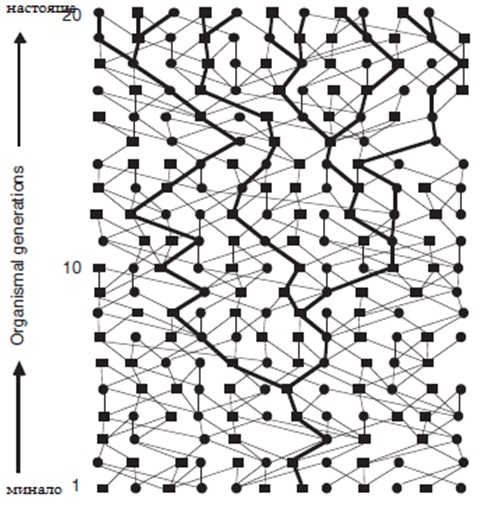 Филогеографията е нова дисциплина, обединяваща няколко дисциплини като генетика, молекулярна билогия, еволюционна биология, зоология, биогеграфия, ботаника и др.
Филогеографията е нова дисциплина, обединяваща няколко дисциплини като генетика, молекулярна билогия, еволюционна биология, зоология, биогеграфия, ботаника и др.