Шведският химик Йонс Якоб Берцелиус (1779 – 1848) е съставил първата таблица на елементите, в която всеки елемент е подреден в съответствие със силата на неговия положителен или отрицателен електрическа заряд. Той е открил, че големината на афинитета между два елемента е пропорционална на разстоянието на което са разделени те в неговата таблица.
Лизозоми (lisosomata)
")
Oткрити са от К. де Дюв в началото на 50-те год. На 20 век.
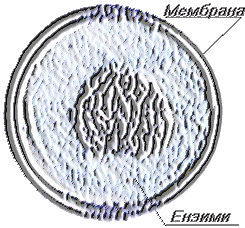
Лизозомите са органиоди и на вътрешната мембранна система на клетката, изпълняваща литични функции те участват в смилането на екзогенните вещества и в елиминирането на отделни вътреклетъчни части (автофагия). Имат кълбовидна, яйцевидна форма и големина от 0,1 до 0,8 µm. Изградени са от гранулирана елекртонноплътна маса, наречена матрикс обграден от двуслойна мембрана, която за разлика от митохондриалната не образува кристи.
Гранулираната вътрешност на йизозомите съдържа голям брой хидролитични ензими, между които значително място заема киселата – фосфатаза, която се смята като маркер на лизозомите. Лизозомите съдържат и други ензими – нуклеази, фосфопротеинфосфатаза и др. Количеството на хидролазите в лизозомите е огромно.
Лизозомите извършват и м-са на апоптолиза – самоунищожение на клетките.
Лизозомите се подразделят на първични и вторични.
Първични лизозоми – Те са изградени от финно гранулиран плътен матрикс. Образуват се от периферните издути части на цистерните на апарата на Голджи. Първичните лозозоми не са активни, защото се съдържат събстрат ( чужд белтък, бактерии и др.)
Вторичните лизозоми са ативни, затова в тях често се срещат полъсмлени или неразтворени остатъци от чужди вещества или собствени вещества от метаболизма на клтката. Вторичните лизозоми се подразделят на :
- Смилателни вакуоли
- Автолизозоми
- Щитоергозоми
- Мултивезикуларни телца
- Телозомни ( остатъчни телца)
Лизозома
ФУНКЦИЯ НА ЛИЗОЗОМИТЕ
Когато „остареят”, част от част от органелите на клетката (митохондрии, едноплазмен ретикулум и др.) се елиминират, като първоначално се ограждат от каналчета на едноплазмения ретикулум, изпълнени смного хидролитични ензими. Тези каналчета, съсдържат хурдрол. зими и огадените цитоплазмени органели образуват автофагозома. Това явление се нарича автофагия. Към тези тени автоагозоми се присъединяват първичните лизозоми и автофагозомите се превръщат в автолизозоми (цитолизозоми). Когато автолизозомите се слеят образуват цитосегрозоми. Когато съдържанието в автолизозомите, респективно в цитосегрезомите се разгради под действието на хидролит. ензими (и след като част пт смлените вещества се използват от клетката ) остават крайни образувания – телолизоми. Веществата от, които се изхвърлят от клетката след преработка.
Лизозомите (автолизозомите и цитосегрезомите) извършват и автолиз (според някой автори понятието е апоптоза) на клетката, ако тя е увредена.
Лизозомите имат и друга функция – модигикация на клетъчни продукти. Наример в клетките на штитпжидната жлеза едноплазмения ретикулум синтезира тироглобулин (белтък предшественик на тиропния хормон) След секрецията на тироглубина от едноплазмения ретикулум, последния се слива с първичните лизозоми, хидлолитичните ензими на които го трансформират в тироиден хормон, който след това се извежда в кръвния поток.
Според съвременните представи лизозомния апарат се е развил като специализирана част на катаболитната система на цитоплазмата с широка функционална пластичност.
ПЕРОКСИЗОМИ
Рази група микро телца е наречена пероксизоми от К. Де Дзв през 1956 г. Те са неголеми вакуоли, обвити с единична мембрана, ограничаваща рганулирания матрикс. В центъра на матрикса има сърцевина. В зоната на сърцевината се намира паракристална структура, състояща се от подредено опаковани фибрили и тръбици.
Пероксизомите са открити при протисти (амеби), при низити гъби (дрожди), при висши растения (ембрионални тъкани, ендосперм), при висши гръбначни животни, главно в черния дроб и бъбреците.
(щракнете изображението, за да го увеличите)
Комплекс на Голджи
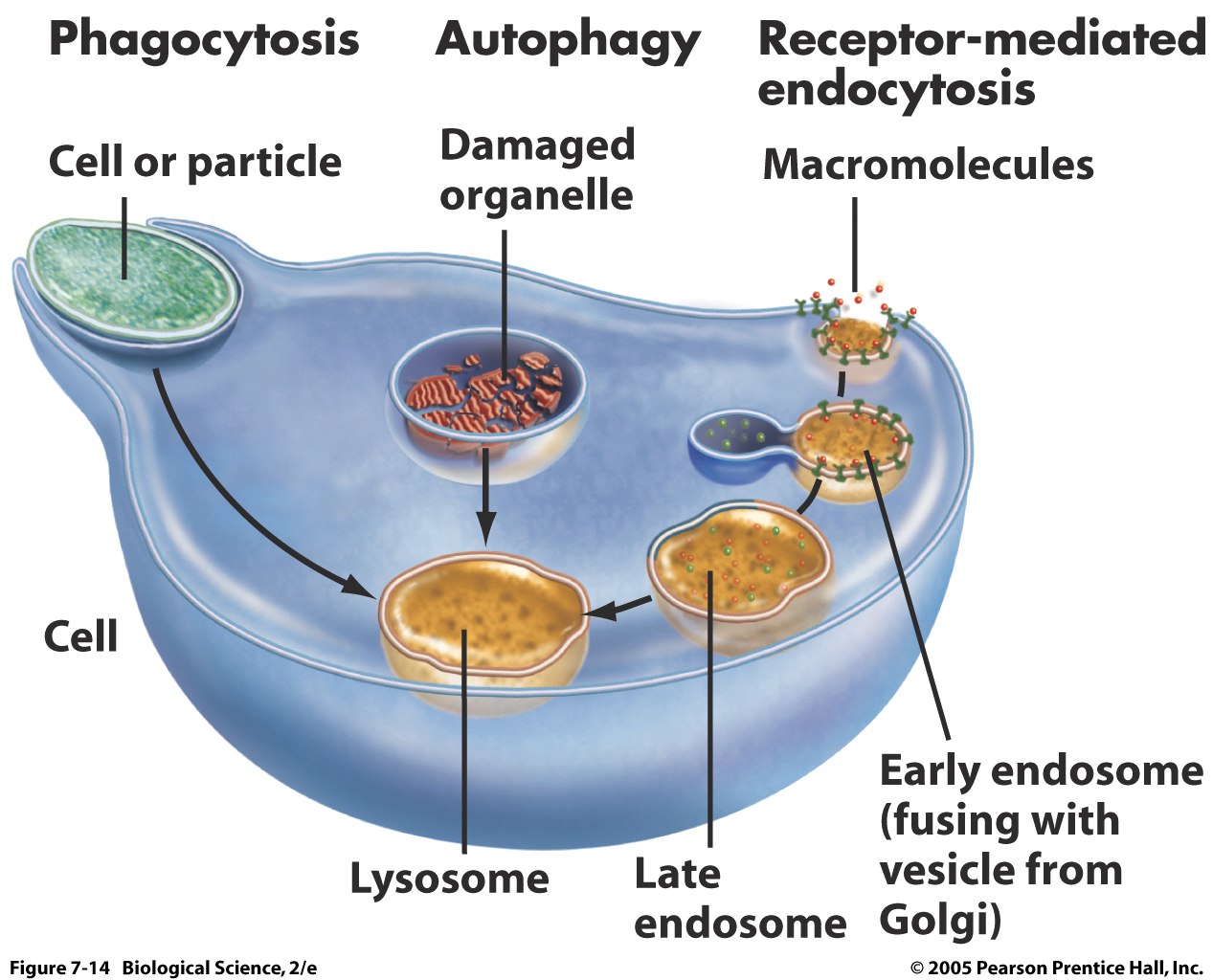
Колмплекса на Голджи е описан през 1898 г. от Камило Голджи. Той го е наблюдавал като мрежест венец около ядрото на нервни клетки.
Обикновено той се намира до ядрото на клетката или около него. Това е свързано с основната му функция – секреторна. Възможно е и промяна на локализацията му в някой клетки във връзка с физиологичните особености на клетката.
Комплекса на Голджи съществува в три нива на организация:
- ЦИСТЕРНА – Сплесната мембранна торбичка с централно плочковидна част и перфорирана периферна зона.
- ДИКЦИОЗОМА – Група от цистери (4-8), наредени в пакет.
- Комплекс на Голджи – Няколко свързани диктиозоми. Броят им варира във връзка с физиологичните потребности на клетката.

Основна структурна единица на комплекса.
Дикциозомата е съставена от плътно разположени една до друга плоски мембранни цистерни, между които има тънък слой цитоплазма. Освен цистерните в зоната на дикциозомата се наблюдават множество вакуоли
АПАРАТ НА ГОЛДЖИ
Комплека на Голджи е полрна структура. Прието е да се различават проксимален (близък, цис) и дистален (далечен – транс) ползси. В секретиращите клетки проксималния край на комплекса е обърнат към ядрото (или едноплазмения ретикумум), а дисталния край към клетъчната мембрана (плазмената). Към проксималния полюс към цистерните на комплекса се доближават малки вакуоли (транзитни) – (транзитните вакуоли идват от едноплазмения ретикулум), а в дисталния край характерзиращи се с по-едрите разширения на цистерните, се образуват големите бакуоли съдържащи продукта на клетъчната секреция ( образувани от комплекса на Голджи)
В клетките отделните дикциозоми магат да бъдат свързани чрез система от цистерни и вакуоли.
Така се формира тримерна мрежа.
! Някой автори вместо понятието дикЦиозома използват понятието дикТиозома.
ФУНКЦИИ НА АПАРАТА НА ГОЛДЖИ !
Апарата на Голджи участва в натрупването и секресията на продуктите синтезирани в едноплазмения ретикулум, участва в тяхното химическо преустройство и узряване, полимеризация на полизахариди и образуване на комплекси от полизахариди (ПЗ) и белтъци (Б) – гликопротеиди; атно на комплекса му липиди(Л) и Б -липопротеиди и др. Освен тези вещества апарата на Голджи натрупва и други вещества – ензими, хормони, vit. Прието е, че в апарата на Голджи се извършва адсорбиране, кондензиране и опаковане на веществата, които се синтезират в гранулирания еноплазмен ретикулум и постъпват чрез транспортни вакуоли в апарата на Голджи. В апарата на голджи се формират и опаковат лизозомите и преоксизомите.
Комплекса на Голджи присъства във всички клетки на еукатиотните организми ( с изключение на еритроцитите при млекопитаещите организми), но не всяка клетка притежава способност за секреция на белтъци, Л и ПЗ (мускулни клетки, еритроцити, покривен епител). Всички те обаче притежават добре развити комплекс на Голджи. Това е свързано с образуването на лизозомите.
НЯКОЙ СПЕЦИФИЧНИ ФОРМИ НА АПАРАТА НА ГОЛДЖИ
ПРИ РАСТИТЕЛНА КЛЕТКА междуклетъчната пластинка, разделяща двете нови, дъщерни клетки след приключване на кариокинезата (делене на ядрото), се обр. от апарата на Голджи. Междуклетъчния плас е продукт на вакуоли от комплекса на Голджи, които изливат съдържанието си в екваториалната плоскост (пектин и хемуцекулата) на делящата се клетка.
Междуклетъчната пластина е изградена от пектин и хемицелулоза.
РАЗВИТИЕ НА АКРОЗОМАТА ПРИ ПРЕРМАТОЗОИДИРЕ
АКРОЗОМАТА Е МЕМБРАННА СТРУКТУРА на сперматозоида, с която той разпознава и свързва сперматозоида към повърхността на яйцеклетката.
Аузиращите от апарата на Голджи вакуоли увеличават повърхността на акрозомата.
(щракнете изображението, за да го увеличите)
Митохондрии

Митохондриите се срещат във всички растителни и животински клетки. Изкючение са едноклетъчните рукариотни организми, живеещи постоянно в анаеробна среда, дифинитивните еритроцити, в роговелите епителни клетки, мутантни дрожди, загубили изцяло или частично митохондрииния си геном. Митохондриите има собствена ДНК и се възпроизвеждат независимо от ядребата ДНК. Наблюденията на митохондриите датират от края на 20в като им са давани различни имена свързани с формата им (митозоми, хондриозоми, биобласти и да. ) В 1898 г. Бехда въвежда името митохондрии, а през 1900 г. Михаелис предлага използването на багрилото. Янус гриин Б, специфично багрещо митохондриите (хистохимичен маркер)
ФОРМА – варира силно дори в пределите на една клетка – специфичниелипсовидни, бъбрековидни, силно източени – нишководни.
РАЗМЕРИ – Дебилината на митохондриите е относително потоянна около 0,5 мкм. Дължината им се коледае от 0,5 до 7-10 мкм при хишководните (силно източените) форми.
Количеството на митохондриите в клетката е различно в зависимост от функцията на клетките.
БРОЯ на митохондриите в клетката зависи от нуждите на самата клетка от енергия и АТФ.
ЛОКАЛИЗАЦИЯТА на митохондриите също е различна, обикновено те се разполагат в тази част на клетката, където има нужда от енергия и АТФ. При недиферинциралихондриите са клетки митохондиите са разположени равномерно в цитоплазмата, а при диферинцираните като например мускулни, митохондриите се намират в близост миофибрилите; в сперматозоидите – завити са като спирала около камшичето и т.н.
СТРУКТУРА
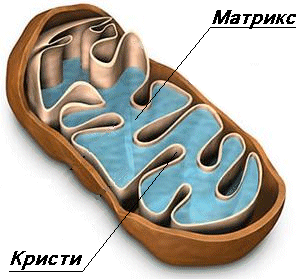
Класическите митохондрии се представят като елипсовидни органоиди, ограничени с две мембрани (двумембранни органоиди), които ги разделят на две пространства – интермембранно пространство и вътрешен матрикс.
Вътрешна митохондриална мембрана – дебелината около 7 nm – отделя съдържанието на органоида от цитоплазмата ( хиалоплазмата). Тя нормално има гладък контур и не е свързано с други мембрани в цитоплазмата притежава неспецифична проницаемост. По статична е, с белтъчно съдържание около 20%. Съдержа порови белтъци – порини през които могат да преминат всички молекули до 5000 D. Други белтъци в тази мембрана включват ензими, свързни с митохондриалния липиден синтез.
Интермрдиарното пространство е изпълнено с хомогенно течно вещвство със слаба електронна плътност. Съдържа предимно нискомолекулни съединения – кофактори и субстрати на биологичното окисление.
Вътрешна митохондриална мембрана – ограничава матрикса (митоплазмата). Тя образува многобройни вгъвания, наречени крисити. Тези нагъвания
(инвагинации) многократно увеличават повърхността на вътрешната мембрана. Криситите са насочени навътре към марикса Вътрешната мембрана се различава от външната. Тя е с висико специфинчна проницаемост, тя е по-гъвкава и по-динамична, с белтъчно съдържание 75%. Това е свързано с електрохимичния градиент на Н , който стимулира АТФ синтезата. Най-общо белтъците на вътрешната мембрана изпълняват три типа функции:
- Узвършват окислителноредукционните реакции в електрон-транспортната верига.
- Синтезират АТФ в матрикса, чрез АТФ – синтези в мембраните.
- Позволяват преминаването на метаболити навън и навътре от матрикса, чрез транспортни белтъци.
МАТРИСК
Висико концентрирана смес от стотици ензими, включително и ензими необходими за цикъла на Кребс. Матриксса има финнозърнест и хомогенен строеж. В него се наблюдават тънки нишки и гранули. Нишките са няколко копия на молекули ДНК а гранулите – митохондриални рибозоми. В матрикса се срещат по-големи гранули които са депо за отлагане на калциеви и магнезиеви соли.
МИТИХОНДРИАЛНА ДНК
Респликацията на митохондриалната ДНК протича независимо от репликацията на ядрената ДНК. Обикновенно репликацията на митохондриалната ДНК става след завършване репликацията на ядрената ДНК. Митохондриалната ДНК е двуверижна, за нея е характерна суперспирализация но има атсъствие на белтъци (при ядрената имаме хистолин). Тя обикновенно се прикрепва вътрешната митохондриална мембрана, т.е за митохондриите с характерна автономия при възпроизвеждането, но по отношение на пълна автономност относно синтезата на пълния набор от белтъци необходими за изпълнение на изпълнение на функиите си, митохондриите често използват белтъци синтезирани от цитоплазмените рибозоми, които постъпват в матрикса и се локализират на специфичните на тях
места.
ФУНКЦИИ НА МИТОХОНДРИИТЕ
- В тях се извършва цикъла на Кребс
- Окислително-фосфолиране ( синтез на АТФ )
- Бета – окисление на мастни киселини
- Елонгиране ( удилжаване ) на мастни киселини
- В митохондриите се отстраняват някой токсични събстанции.
Митохондриите са сързани главно с енергитичния метаболизъм на еукариотната клетка клетката – синтез на АТФ. Стратегията на цикъла на Кребс е същата с тази на гликозата ( извършва не при анаеробни условия в цитоплазмата),а именно: Разграждане на голени въглеводородни молекули с отделяне на енергия по формата на АТФ или под формата на НАДН2 и ФАДН2
ОКИСЛИТЕЛНО – ФОСФОРИЛИРАНЕ – Водорода от НАДН2 и ФАДН2 се подава по веригата от най-малко пет междинни субстанции [Тази верига се нарича електрон-транспортна верига (дихателна верига)]. Този водород по тази верига достига до молекулата на О2 и се образува Н2О. Всяко пренасяне на Н редуцира (приема е-) на донорната молекула.
Преноса на Н в началото на веригата, а в края на е- създават електроден градиент който е причина за фосфолиринато на АДФ до АТФ. В този процес участват оксизолите в чиито главички е разположен АТФ – цинтетазния комплекс
Я-ОКИСЛЕНИЕ – Свободни мастни киселини се отркиват в клетката само в следи, защото са силно токсични. В цитоплазмата масните кислелини се свързват с глицерол в моноди и триглицериди и фосфолипиди. Те се хидлолизират в цитокл. и се транспортират в митохондриите кдето се окисляват напълно
(Я-ОКИСЛЕНИЕ )
ЕЛОНГОРАНЕ НА МАСТНИ КИСЕЛИНИ – Най-общо мастните киселини се синтезират в агранулирания едноплазмен ретикулум но в митохондриите също има ензими катализаращи елонгирането на палмитинова и др. наситени мастни киселини.
Хидразин

Частично окисление на амоняк (NH3) води до получаване на хидразин.

2NH3 NaOCl –> N2H4 H2O NaOH
Хидразинът представлява безцветна, димяща на въздуха течност (т.т. 1,4°С ; т.к. 113,5°С )
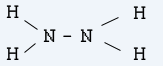
Хидразинът и водните му разтвори се използват като редуктори при което се окислява до азот.
HO2 CH3N2H2 –>
Анализ на месинг

Количественото определяне на съставките на сплавите заема значителен дял от приложния анализ. То е от голямо значение за металургията и за металознанието.
Анализът на месинг е един от най-характерните анализи на сплави. Както е известно, съставът на месинговата сплав включва Cu в преобладаващата част, Zn също в значително количество, а в малки количества— от следи до 1—2% — Pb, Sn и Fe.
Пробата се подготвя под формата на стружки. За изходно количество се взема около 0,5 g и се претегля с точност до четвъртия знак. За разтварянето й е достатъчно да се залее в бехерова чаша с вместимост от 150 до 200 cm3 с около 10 cm3 разр. HNO3(2 : 1). Буйно протичащата реакция води до опасност от изпръскване. Ето защо чашата се покрива с часовниково стъкло. Ако останат не разпаднати стружки, чашата леко се нагрява. Доба вят се 60 cm3 дестилирана вода, нагрята до кипене, и внимателно се смиват стените на чашата с пръскалката, като пръските от получавания разтвор се задържат от покриващото часовниково стъкло. За да приключи реакцията, необходимо е допълнително леко нагряване в продължение наполовин час. При обработването на сплавта с HNO3 калаят преминава в метакалаена киселина:
3Sn 4HNO3 H2O –>ЗH2SnO3 4NO.
Тя пада като утайка, филтрува се през плътен количествен филтър (предпочита се синя лента). За да се освободи утайката от разтворимите примеси, промива се с гореща вода. В нея предварително се разтваря малко количество от летливия електролит NH4NO3.
Промитата утайка се суши при 118°С. След това се нагрява в плитък порцеланов тигел, като се осигуряват окислителни условия на накаляването. Протича следната реакция:
H2SnO3=SnO2 H2 O
Когато се достигне постоянна маса, утайката се тегли като SNO2 и се изчислява в проценти спрямо изходната маса на пробата.
Следващият метал, който се отлъчва, е Pb. Когато се прилага тегловният метод, за подходяща форма на утаяване се избира PbSO4. Намиращият се във филтрата Pb(NO) 3) 2 се превръща в PbSO4, като се действува със H2SO4:
Pb(NO3)2 H2SO4=PbSO4 2HNO3
Тъй като NO3 силно влияе на разтворимостта на PbSO4, за да не се компрометира резултатът, налага се HNO3 изцяло да се отстрани от филтрата. За целта към него се добавят 4 cm3 конц. H2SO4 и се изпарява в порцеланово блюдо, докато започнат да се отделят гъсти пари от SO3– Препоръчва се поне веднаж да се прекъсне нагряването, за да се свалят със струйка вода отложените по стените на блюдото соли.
Когато изпаряването привърши, остатъкът в блюдото се залива с 25 cm3 студена вода. Получената кора от соли се раздробява със стъклена пръчка. Подлага се отново на нагряване до кипване. С това се цели да се отделят разтворимите във вода сулфати от кристалите на утайката на PbSO4. Доливат се отново 50 cm3 вода и след като утайката се охлади в продължение на един час, се филтрува през предварително изсушен и претеглен филтър-тигел. Филтруваната утайка първоначално се промива с разредена H2SO4, а след това с алкохол. С това по-пълно се ограничава разтворимостта на PbSO4 в промивайте води. Промитият филтър-тигел се суши при 200° С до постоянна маса.
Определянето на Pb може да се извърши и ускорено но електроана-литиченпът. Това обаче е свързано с употребата на скъпи платинови електроди.
Третият метал, който се отлъчва, е медта. Във филтрата, получен от отделянето на PbSO4, той се намира като CuSO4 и може да се определи както тегловно, така и електроаналитично.
При тегловното определяне се спазва следната последователност.
Филтратът се концентрира на водна баня в порцеланово блюдо до обем от около 150 cm3. Прехвърля се количествено в конична колба. Добавят се 15 cm3 конц. H2SO4. Когато температурата му спадне на 70°С, през него се пропуска бавно струя от чист H2S. Получената утайка трябва да се филтрува незабавно поради опасността от окисляването на CuS до CuSO4 под действието на атмосферния кислород. Използува се смукално шише е филтър бяла лента. Измива се с 4%-ен разтвор на оцетна киселина, наситена със H2S. С това се предотвратява пептизацията на утайката от CuS. Едно от важните изисквания както при филтруването, така и при промиването е филтърът да бъде винаги запълнен с разтвор. С това се препятства окислителното действие на атмосферата, в резултат, на което CuS частично преминава в разтворимия CuSO4.
Утайката се суши, след което се отделя количествено от филтъра върхубяла гланцова хартия. Така обработеният филтър се изгаря в предварително накален и претеглен тигел на Розе. След това се прибавя и утайката. Накаляването се извършва за късо време при достатъчен достъпна въздух. По този начин филтърните влакна, увлечени в утайката, изгарят на-пълно. След като утайката се охлади до стайна температура, в тигела се насипва химически чиста сяра така, че да покрие утайката. Поставя се и капакът и неговата луличка се свързва с Киповия апарат. Произвежданият в него водород предварително се прекарва през промивно шише, съдържащо алкализиран разтвор от KMnO4. Целта е да се задържат минималните количества арсеноводород, които се отделят при получаването на водород в Киповия апарат. Присъствието му в тигела би повлияло на точността на резултата.
След като водородната струя изпъди въздуха от тигела на Розе, неговото дъно се нагрява с горелка до слабо червена жар. При това сярата, която сублимира, и водородът се запалват на изхода на тигела. Създадените условия, както лесно можем да се убедим, не допускат утайката от CuS да се окисли до CuSO4. След нагряване от 10 min при усилена струя се спира горелката. Подаването на H2 спира след още 2—3 min. Тигелът се темперира в ексикатор и претегля. След това нагряването при описаните условия продължава, докато се получи постоянна маса.
Утайката е под формата на Cu2Sпоради излишъка от химически чиста S, с която се обработва филтруваната утайка от CuS. Ето защо при изчисленията се взима под внимание този факт.
Значително по-лесно се извършва електро аналитичното определяне. Филтратът се концентрира до около 100 cm3. Към него се добавя 1 cm3 конц. HNO3 и се електро анализира.
Предпоследният метал от сплавта, който се отлъчва, е желязото. Неговото определяне се извършва по познатия тегловен начин, и то направо катоFe(OH) 3, ако преди това медта е отлъчена електро аналитично. Ако е следван тегловният метод за определяне на медта като Cu2S, се извършва следната предварителна обработка на филтрата. Отстранява се съдържащият се в него H2S (останал от насищането при предишното отлъчване). Това се постига с продължително нагряване на филтрата. В противен случай Н28 предизвиква примесване в утайката и на FeS, което понижава точността на резултата.
За да може Fe3 да паднат като утайка от Fe(OH) 3, предварително се окисляват с 2—3 капки бромна вода или конц. HNO3. Следва утаяване с 2n NH3. Разликата от познатия тегловен метод за определяне на Ре е. в употребата на значителен излишък от NH3. С това се предотвратява съвместното утаяване и на Zn като Zn(OH) 2. В останалия си ход определението следва познатия начин на работа.
Последният метал в месинга, който трябва да се отлъчи, е Zn. Филтратътс промивните води от предишното определение се изпарява до сухо в порцеланово блюдо, което се загрява на водна баня. За да се отстрани нежеланият излишък от летливите амониеви соли, остатъкът върху блюдото се накалява леко. След това се разтваря във вода, подкиселена слабо съсHCl. От така подготвения разтвор Zn2 -йони се утаяват като ZnNH4PO4. Това се постига, като средата предварително се неутрализира с NH3.За неутралната точка се следи с няколко капки от индикатора метилрот. След като разтворът се нагрее до кипване, прибавят се 20 cm3 20%-ен разтвор от (NH4)2HPO4. Той трябва да бъде прясно приготвен. За да могат да се превърнат наличните Zn2 йони изцяло, в добре кристализиращия ZnNH4PO4,кипенето продължава върху водна баня в продължение на половин час.Внимава се утаяването да не се извършва в кисела среда. Ето защо не трябва да се променя жълтият цвят на индикатора метилрот. За да може кристалната утайка да узрее добре, изчаква се около един час. Филтрувасе през количествен филтър бяла лента. Поради склонността на Cl– -йони, да се задържат в кристалите на ZnNH4PO4 утайката се промива 1%-ен разтвор на утаителя. Когато в промивните води се констатира с помощта на AgNO3 отсъствието на Cl– -йони, промиването продължава с 50%-ен алкохолен разтвор. Така се отстранява нереагиралият (HN4)2HPO4.
Промитата утайка се суши.. Снема се от филтъра върху черна гланцова хартия. Филтърът се изгаря в предварително накален и претеглен тигел. Съвместното изгаряне и накаляване на утайката с филтъра може да доведат до включване в получаваните кристали от Zn2Р2О7 на недоизгорели въглеродни частици. А това не позволява тяхното отстранявана при температура на накаляването и намалява точността на резултата.
След като филтърът се изгори в тигела, прибавя се количествено и утайката и се подлага на накаляване до постоянна маса. Утайката се превръща, в Zn2P2О7 по уравнението
2ZnNH4РО4==Zn2 Р2О7 2NHз Н2О
На тази стехиометрична основа се изчислява съдържанието на Zn в месинга.
Като ориентировъчни за правилността на резултата от анализа могат да служат следните гранични, стойности, с които отделните метали участвуват в състава на различните видове месинг: Sn—0 до 1%; Pb—-от 0до 2%; Cu—от 55 до 72%; Fe—от 0 до 1%; 2п—от 25 до 45%.
Анализ на бронз

Както е известно, бронзът е също медна сплав, в която за разлика от месинга вторият по значение метал е калаят. В нея може да се съдържа и фосфор. Тези разлики обуславят и някои особености в хода на анализа.
Поради много по-голямото количество калай, който се съдържа в бронза(до 20% спрямо 1% максимум при месинга), при отлъчването, му се получава обемиста утайка от метакалаена киселина. Тя е онечистена от хидроокисите на медта, а също така въпреки и по-слабо — от хидроокиситена цинка, оловото и фосфора. Количеството им не може да се пренебрегне.В накалената утайка от SnО2 те се примесват като окиси и компрометират резултата.
За да се отстранят, накалената и претеглена утайка от SnО2 се стапя със смес от Na2СО3 и S. В стопилката Sn се съдържа като Ка2SnS3, Р — като Na3РО4, докато СuО, ZnО и PbО остават непроменени. За да се отделят, стопилката се изварява с вода, след което полученият неразтворим остатък се фитрува. След промиването му с гореща вода се накалява до постоянна маса и се тегли. Получената стойност се приспада от първоначалното тегло на 8пО2 (заедно с примесите). Така се намира количеството на SnО2 и Р2О5.
За да не се загубят Pb, Zn и Сu за по-нататъшния ход на анализа, отстраненият остатък се разтваря в няколко капки конц. HNO3 и се добавя към филтрата, получен при отлъчването на Sn.
Ако се изследва фосфорен бронз, разработва се втора проба. Изходното количество на пробата е по-голямо — 2 g. Отлъчената по описания начин утайка от хидроокисите на Sn и Р се отделя количествено от филтъра. След накаляването и се смесва в тигел с капаче с трикратно повече KCN. Тигелчето се нагрява, докато получената стопилка престои в течно състояние 20 минути. В резултат на протичащите взаимодействия се отделя метален калай, докато Р2О5 от накалената утайка минава в разтворимия К3РО4. За да може това съединение да се извлече от втвърдената стопилка, тя се изварява с вода. Неразтворимият остатък (калаят) се филтрува и промива. Полученият филтрат се подкиселява със солна киселина. Намиращите се в остатъка минимални количества Сu2 и Sn2 се утаяват чрез насищане на разтвора със Н28. Падналата утайка от СuS иSnS2 се отстранява чрез филтруване. Самият филтрат се нагрява на водна баня, докато се изпъди останалият в него Н2S. Съдържащите се в разтвора РО43- -йони се определят тегловно като МgNН4РО4. За целта се прибавят 1 g МgСl2 и 2—3 g NH4Сl. Разтворът се подкиселява със НСl. По-нататък, утайката се разработва, както при тегловното определяне на Zn.
Изчисленото по стехиометричен път съдържание на Р2О5 се изважда от общата маса SnО2 Р2О5, с което се определя количеството на SnО2. Посредством него се изчислява и процентното съдържание на медта в бронза.
Количественото определяне на останалите метали следва същия ход, както и при анализа на месинга,
Правдоподобността на резултата може да се прецени ориентировъчно последните гранични данни за състава на отделните разновидности бронз: Cu— от 82 до 96%; Zn — ‘от 0 до 8%; Pb — от 0 до 3%; Sn — от 4 до 20%; P—от 0 до 1%.
Серният диоксид

Серният диоксид е безцветен газ с характерна миризма, т.т. -72,70С и т.к. -100 С. Получава се при горене на сяра, метални сулфиди или сероводород на въздуха или от киселина и сулфит или хидро генсулфит. Силен редуктор,особено във воден разтвор. Разтваря се във вода и дава хидрат на газа,като разтворът се отнася като киселина /серниста киселина/. Използва се за производството на SO3 за получаване на сярна киселина.
Белтък

Белтък (или протеин) е колективен термин за биологически важни макро молекули, изградени чрез поликондензирането на отделни аминокиселини.
Белтъците са един от основните градивните компоненти на живите клетки, както и на вирусните частици. Притежават сложна пространствена структура, и изпълняват разнообразни биологични функции — от типично структурни, защитни, траспортни до каталитични и регулаторни.
Цялата информация необходима за изграждането на белтъчната молекула е кодирана в ДНК. Посредством процес на точно презаписване на информацията в РНК (транскрипция) и превеждане от полинуклеотидна в аминокиселинна последователност (транслация) е възможно синтезира определен белтък, притежаващ конкретна биологична роля.
За първи път през 1789 г. френският химик Антоан Фуркроа прави разграничение между белтъците и другите известни органични субстанции.Класифицира познатите дотогава албумин, фибрин и желатин като самостоятелни съединения с животински произход. От тази начална стъпка до формулирането на концепция за това, какво представляват белтъците, е извървян дълъг път, минаващ през грубо характеризиране на химичния състав, опити за клинично приложение на белтъчни екстракти, до все по-прецизното дефиниране на структурата и функциите на всеки отделен белтък.
Самото понятие е въведено през 1838 година, когато в писмо от 10 юли,изпратено от Йонс Якоб Берцелиус до Герардус Йоханес Мулдер, за първи път е употребено названието „протеин” с акцент върху първостепенното и първичното (от гръцки πρωτειοξ — първична нишка). Точният цитат от писмото е:
«Le nom protйine que je vous propose pour l’oxyde organique de lafibrine et de l’albumine, je voulais le dйriver de πρωτειοξ, parcequ’il paraоt кtre la substance primitive ou principale de la nutritionanimale.»
Основният компонент на всички белтъци са аминокиселини, подредени последователно в дълги полимерни вериги. Връзката между отделните аминокиселини е амидна, но поради някой свой особености е наречена със специфично име – пептидна връзка. Затова и самите полимелни вериги се означават като полипептидни. Организацията в пространството на полипептидните вериги е изключително сложна и характерна за всеки отделен белтък. За прегледност и по-лесна ориентация, пространствената структура на белтъците е разделена на отделни нива:
#първично ниво (примерна структура) – представлява точната последователност на свързване на отделните аминокиселинни остатъци.
#вторично ниво (секундерна структура) – това са локални нагъвания на основния скелет на полипептидната верига, притежаващи известна периодичност. Всяка възпроизводима и характерна форма се означава като отделен тип вторична структура, като например алфа-спирала,бета-листовидна структура, бета- и гама-завой, както и липса на порядък.
#третично ниво (терциерна структура) – пълното нагъване на полипептидната верига в пространството, с всички възможни близки и далечни взаимодействия. Това ниво дава престава за цялостната форма на белтъчната молекула, както и за връзките и отношенията между отделните вторични структури.
#четвъртично ниво (кватернерна структура) – това е отново цялостна пространствена форма, но на асоциирани не ковалентно белтъчни молекули. Всяка отделна белтъчна молекула, притежаваща характерна, завършена, третична стурктура се означава като суб-единица, а цялостният конгломерат – като олигомер. Не всички белтъци притежават четвъртична структура, тъй като повечето са изградени от една полипептидна верига,а четвъртично ниво предполага наличието на поне две. Типичен пример за белтък с четвъртична структура е хемоглобина.
Всички тези нива дават престава за строежа на отделната белтъчна молекула, но по същество са стационарни описания на структурата.Белтъците в разтвор, както нормално съществуват в живите клетки, търпят вариации в структурата си, тъй като са подложени на редица въздействия.Тези вариации могат да бъдат малки отклонения – наричани още „дихателни движения“, но могат и да се отразят съществено на структурата и сериозно да я видоизменят.
Пространствената структурата на белтъка се нарича конформация, а всички изменения, независимо дали са съществени или не, се означават като конформационни промени.
Всеки белтък притежава специфична функция, като практически няма процес в живите организми, който да не зависи от конкретен протеин. Функциите могат да са най-разнообразни, но формално могат да се обединят в няколко основни:
структурна функция – белтъци, участващи в архитектурата на клетките, поддържащи определена форма и вътрешна организация.
регулаторна функция – това са всички белтъчни системи,които регулират процесите в клетката – белтъци от сигналните пътища,фактори на транскрипцията и транслацията, и не на последно място хормоните.
защитна функция – белтъци, осигуряващи чисто механични защитни бариери. При висшите животински организми има специални системи за защита срещу определени патогени, като белтъците отговорни в най-голяма степен за това се наричат антитела.
каталитична функция – протичането на почти всяка химична реакция в организма, се дължи на определен белтък, който в този случай се означава като ензим.
транспортна функция – това са белтъци, пренасящи определени молекули през клетъчни мембрани, или транспортиращи вещества на големи разстояния – кислород пренасящите протеини, като хемоглобин, миоглобин и други.
двигателна функция – всички мускули при животните, савсъщност функция на съответни белтъци, като например актин и миозин. Често такива белтъци се означават като „молекулни мотори“. Участват във всяко движение – било на клетъчни органели в рамките на клетката или на целия организъм в пространството.
Планиране и развитие в професионалната кариера – етапи на мотивацията за постижения


За едно успешно развитие на дадена организация е необходимо развитието и на служителите в нея да бъде от съществено значение. Не един път е спомената тъй важната зависимост между двете страни. Наименованието на колектива „движещата сила на всяка организация” не е дадено случайно. Мотивацията за действие на тази движеща сила не зависи само от потребностите, мотивите, стимулите, а и от формираните крайни дългосрочни цели, които ги обединяват.
Със сигурност кариерата на всеки отделен член на колектива не е просто факт. За нейното развитие и просперитет изключително важна роля изиграва самата личност, нивото на нейната мотивация за действие, работната и организационна среда- предоставените от нея възможности. Кариерата на всеки човек преминава през серия от фази, като всяка една от тях може да бъде повлияна от известни фактори обвързани с мотивацията за постижения. Намирането на адекватно съответствие между индивидуалните потребности, способности, мотиви, и организационни възможности не е нещо, което просто се случва. По този въпрос главно се разглеждат основните две страни, от които изцяло зависи очертаването на кариерният път на отделния кадър – нагласата на организацията,възможностите, мотивацията в лицето на кадъра.
За организациите в по- големия процент от случаите е необходимо да осъзнават потребностите на служителите си, като по този начин е на лице реалността за преценка и подбор на кадрите в персонала. Главната цел за тази равносметка е използване на най- способните и просперитетни служители по най- добрия начин касаещ и двете страни.
От друга страна възможностите на тези служители обикновено са обвързани със завишена мотивация и амбиция за професионално и личностно израстване, което обвързва организацията с предоставяне на ясна картина за наличието на очакваните възможности в бъдеще. Споделянето на информация и разбирането на „етапа на кариерата” са от съществено значение и в голяма степен играят роля на силен стимул, непрекъснат стремеж за планиране на кариерата и усилия целящи нейното развитие.
Етапи на кариерата
Идеята затова, че хората преминават през различни етапи на своето личностно и професионално развитие е всеизвестна и широко призната. Един от най- основните варианти включва четири етапа:
– етап на пред-работата (обучение )
– първоначален трудов етап (местене от работно място на друго, доказване на личностните качества и възможности)
– стабилен трудов етап (затвърждаване на позициите и поддържане на изградения образ)
– етап на пенсионирането (равносметка за изтеклия трудов стаж и личната и професионална удовлетвореност или неудовлетвореност от него) .
В повечето случаи амбициозните хора с дългосрочна цел кариерно развитие се подготвят за него като преминават през определен курс на обучение зависещ от редица фактори като стремежи, нагласи, очаквания. Той може да бъде изключително кратък, както и изключително продължителен поради тази причина. След това постъпват на работа в първата организация с подобна на тяхната амбиция за постижения насоченост и област сфера, която може да се окаже както единствената (в случай че им предостави желания от тях краен резултат),така и една от няколкото поредни (поради не реализация, при наличие на отрицателно стекли се обстоятелства или поради други причини).
Едно от най-важните решения, които човек се налага да вземе е избора на работа,кариера. Сериозността на този избор е изцяло насочена към първоначалната мисъл за постоянството при работа и семейство с продължителност, касаеща целия живот на човек. Поради тази причина разнообразността от занимания и професии е изключително широка и зависеща в много голяма степен от типа личност и работната среда. При грешен избор на занимание не може да се очаква силна мотивация за постижения, тъй като желанието за развитие и растеж в сфера в която няма реален интерес е немислимо..
Холанд смята, че мотивацията и продуктивността на всеки един от тези типове личност се влияе от насочеността към типа работа и нейната връзка с личностната характеристика (качества, нагласа, предразположеност). При несъответствие между двата фактора в по- големия процент от случаите се наблюдава и отражение в нивото на мотивация за постижения и продуктивност на работното място. John L. Holland (1966).
Това твърдение е на лице при Холанд, който смята, че се обособяват шест обобщени личностни типа влияещи на мотивацията за работа и постижения в сферата на кариерноторазвитие
1. реалистичен – Този тип личност е с насоченост на интересите към манипулация на машини и съображения. Отличава се с атлетични или механични способности.
2. изследователски – Този тип личност се отличава с изследователски, аналитичен, любознателен характер. Мотивацията за постижения при него е свързана с желанието за постоянно развитие от мисловен, научен тип. Всяко едно действие свързано с работата му се отличава с изключителна прецизност.
3. артистичен – Този тип личност е експресивен, интроспективен. Това са хора притежаващи артистични, художествени, иновативни, интуативни способности. Главното което използват в работата си и от което зависи нивото на мотивация за постижения при тях е креативността, представата – усещането за крайния резултат, (музата), въображението.
4. социален – За този тип личност са характерни контактността, нагласата и желанието за постоянна комуникация и контакт с хората. Главното от което зависи мотивацията за постижения е мисълта за срещите с хората и помощта която ще им бъде оказана.
5. предприемчив – Този човек изпитва удовлетвореност от влиянието си над другите. Мотивацията за постижения при хора с такава насоченост е характерна с нивото на влияние, убеждение, управление на човешкото поведение.
6. конвенционален – Този тип е характерен за работниците в административния сектор, тъй като мотивацията и прецизността са насочени към спокойния стереотипен, неангажиращ с управление, влияние, въображение, изобретателност начин на работа.
Процес на планиране на кариерата
Хората и организациите започват да осъзнават все повече важността на планиране на кариерата от гледна точка на мотивацията.
Практиката на планиране на кариерата в повечето от случаите включва намирането на съответствие между аспирациите на човека за кариера и възможностите предоставени му от организацията. Процесът на планиране на кариерното развитие, начертаването на кариерен път, неговото успешно използване поставят еднаква отговорност върху индивида и организацията. Това, което зависи от индивида е личната равносметка относно възможности, компетенции, начини за развитието и реализацията им. Организацията от друга страна като институция идентифицира потребностите и възможностите на индивида и чрез благоприятен мениджмънт осигурява необходимото обучение, реализация на служителите си.
Литература:
[1] John L. Holland ThePsychology of Vocational Choice: A Theory of Personality Types and ModelEnvironments in 1966.
John L.Holland Journalof Counseling Psychology in 1959
Gary D.Gottfredson John L.Holland’s Contributions to Vocational Psychology:
A Reviewand Evaluation
[1] Douglas T. Hall(1976). Careers in Organizations. Santa Monica. CA: Goodyear Publishers, p.4.
Walter Kiechel III (1983). TheNeglected Art of Career Planning. Fortune, June 27, pp. 153-155
Източник: nauka.bg