Една звезда се ражда в резултат от гравитационно свиване на чудовищен газов облак. Подобни облаци изпълват част от нашата Галактика и се появяват (в телескопите) като тъмни слабо тлеещи мъглявини, които затъмняват намиращите се зад тях звезди. Където и да съществуват тези облаци, от тях се раждат звезди
.М16
Междузвездните облаци се състоят главно от водород. На всеки кубичен сантиметър се падат по около осем атома и една молекула. Освен това има и следи от други молекули, като при това част от последните са органични. Протозвездите възникват в някои от локалните струпвания на вещество, като във вътрешността на облака се наброяват до десетки хиляди.Локалните струпвания на материя привличат към себе си все повече материя благодарение на гравитационното привличане. Докато атомите имолекулите падат свободно към тези струпвания, те набират скорост,блъскат се една в друга и придобиват тази хаотична форма на движение,която наричаме топлина. Следователно гравитационното свиване на газаповишава топлината му. Разредеността на междузвездния газ е толкова голяма, че процесът на образуване на звездите продължава около 10 000000 години.
В зависимост от масата на газа, който се свива, звездата маже да е мъртвородена, нормална или нестабилна.
Слънце
Полезна мярка е масата на Слънцето – 1,99×10і° кг която бележим със символа М☼. Ако масата е по – малка от около 0,1М☼ гравитационното налягане е недостатъчно за да се достигне температурата 10 000 000 К, която е необходима за термоядрен синтез на хелий от водородни ядра. Ако масата превишава 100☼, гравитационният натиск е така силен, че след като веднъж е започнал термоядрен синтез, във вътрешността на звездата се поражда радиационно налягане което превишава нормалното налягане на газа. При тези условия звездата става хидродинамично неустоичива и трябва да изхвърли взривно материя, за да се нормализира. Следователно нормалните протозвезди имат маса между 0,1М☼ и 100М☼. След като температурата в центъра на звездата се повиши до 10 000 000 К, термоядрената енергия, получена при синтеза на хелиевите ядра от протони, прекъсва по-нататъшния процес на гравитационно свиване. В такъв случай казваме че звездата принадлежи на главната серия. Тогава тя се характеризира с ядро от ”горящ водород”, със стабилен радиус и с излъчвана енергия , равна на енергията която се освобождава при превръщането на водорода в хелий. Такава звезда е нашето Слънце.
Колко дълго една звезда остава в главна серия, зависи от това колко време е необходимо за изразходване на водорода в ядрото й. Една звезда с маса 15 М☼ ще оставе в главна серия около 10 000 000 години, което е около 460 пъти по малко от възрастта на земното кълбо. Това, че ние виждаме такива звезди по небето, е доказателство за това, че в нашата галактика продължават да се раждат звезди. От друга страна, очаква се, че Слънцето ще остане в главна серия около 8 000 000 000 години. Ако Земята и Слънцето са на една възраст, то това означава, че Слънцето трябва да прекара спокойно още 3 000 000 000 години върху главната серия. Ако обаче Слънцето се е сдобило с планетната си система , след като е било вече образувано, както твърдят някои теории, то няма никакви данни за това, колко дълго ще остане то в сегашното си състояние.
Какво става когато водородът в звездното ядро бъде изразходван? Смята се, че щом като при това се увеличава количеството на хелия, то звездата се загрява още повече и става още по ярка. На такъв етап хелият напълно заменя водорода във вътрешността на звездата. Подробностите в поведението на звездата след достигането на тази критична точка силно зависят от това, колко голямо е смесването на газовете. Това от своя страна зависи от масата – смесването е толкова по – голямо колкото по – голяма е масата. Изобщо казано, след като веднъж е образувано хелиевото ядро на звездата, не могат да протичат вече никакви ядрени реакции. Ето защо е необходимо ядрото да се свие още повече, за да се поддържа достатъчно висока температура в обвивката му, която да позволява на останалия в нея водород да гори. След като хелиевото ядро достигне някаква критична маса, то става механично неустойчиво и бързо се свива и се нагрява. Едновременно с това външните слоеве се разширяват и повърхността се охлажда. Звездата се превръща в ”червен гигант” с радиус от петдесет до сто пъти по голям от слънчевия.
Червен гигант
При стократно разширение повърхността на Слънцето ще надхвърли орбитата на Венера и ще достигне до половината от сегашното си разстояние до Земята. Нашата планета ще стане необитаема далеч преди достигането на състоянието на червен гигант, горе долу по времето, когато значително ще се изчерпи водорода в центъра на Слънцето.
Един червен гигант в никой случай не е удобен съсед. Дълбоко в недрата му е заложена потенциална хелиева бомба. Ако масата на звездата превищава 0.4 М☼, в резултат на гравитационното свиване температурата в центъра ще се повиши и ще достигне 100 000 000 К, когато хелият ще се запали /в ядрен смисъл/, като започне да произвежда въглерод. Когато стане това, звездата ще бъде разтърсвана от конвулсии, по време на които в междузвездното пространство може да бъде изхвърлено значително количество материя. След изчерпването на хелия във вътрешността на звездата се образува въглеродно ядро и тя претърпява нова фаза в еволюцията си. Тя отново става ”червен гигант”, но с много по голяма яркост отпреди. Ако е достатъчно масивна, звездата може да достигне до температури, при които може да ”изгаря” въглерода. При това отново се изхвърля взривна материя в междузвездното пространство. Цикълът би могъл да се повтаря и с други термоядрени горива, докато не се образува желязно ядро. От там на сетне не е възможен друг източник на енергия освен чрез гравитационното свиване. Счита се, че в крайна сметка при тимпиратури от около 7 000 000 000 К протичат разннобразни ядрени процеси и взаимодействия между елементарните частици. Те или поглъщат енергия или я предават на неутрината, които свободно напускат звездата. Полученото по този начин охлаждане води до катастрофален колапс. Това от своя страна ще доведе до бързо нагряване на външния слой, а това пък ще накара водорода и хелия останали в него да участват на ново в термоядрен синтез, поради което голяма част от звездата ще се разлети на части.
Свръхнова
Същевременно в хода на този процес се раждат редица елементи, по – тежки от желязото. Счита се, че това е пътя по които се раждат свръхновите звезди и по – тежките от желязото химични елементи.
След като изразходва всичкото термоядрено гориво, звездата или това, което е останало от нея в процеса на еволюцията й се охлажда. Ако масата й е по – малка от 1.4 М☼ тя се превръща в ”бяло джудже”, което постепенно изстива до ”черно джудже” с диаметър 100 пъти по – малък от този на Слънцето.
Бели джуджета
Насоченото навън налягане в звездите, чиято маса е по – голяма от 1.4 М☼ не е достатъчно голямо, за да противостои на гравитацията. Поради това такива звезди се свиват. Все пак ако масата е по – малка от 2 М☼ това свиване стига до стабилно крайно положение, когато плътноста се изравни с ядрената (около 100 000 kg.m‾ і)..
Неутронна звезда
При тези условия електроните се натикват в атомните ядра и се включват в неутроните. Крайната фаза в еволюцията в този случай е неутронна звезда, чиито диаметър е около 100 км.
Твърде правдоподобно е, че тези обекти са идентични с пулсарите – малки тела които излъчват радио импулси с период от 30 милисекунди. Ако масата на звездата превишава 2 М☼, нищо не може да предотврати свиването й до безкрай тоест, при което се раждат така наречените ”черни дупки” – обекти, чието гравитационно поле е така мощно, че дори светлината не може да се отскубне от него.
Черна дупка
Колкото по – масивна е една звезда толкова по – бързо еволюира тя. Това води до интересни последици в звездите, които са гравитационно свързани по между си в двойна система. По – масивната компонента еволюира по – бързо и по – скоро от своя малък партньор достига фазата на червен гигант. При разширението на ”червеният гигант” е възможно да се получи ефектен пренос на материя от по – масивната към по – малко масивната звезда.
Щом Слънцето не е член на двойна звездна система и има скромна маса, би следвало да се очаква, че в бъдеще му е отредено да доживее спокойно дните си като ”бяло джудже”. Това е доста спокойна съдба предвит на това, което изобщо може да се случи. Няма съмнение обаче, че условията за живот на Земята ще станат неблагоприятни още щом Слънцето започне да еволюира от стабилното си поведение на член от главната серия към поведението на ”червен гигант”.
Вероятно това ще се случи след много милиарди години. Време достатъчно да се развият и умрат десетки хиляди технологични цивилизации. Време достатъчно, за да се прокара пътя за отстъпление на живота от Земята. Но ние не можем да бъдем сигурни в това. Смятаме, че Земята и Слънчевата система са възникнали преди
4 600 000 000 години. Дали и Слънцето е възникнало по същото време? Дали Слънцето и планетите са се образували от една и съща първична мъглявина по едно и също време? Ние си даваме сметка за това, че огромното количество материя, която наричаме Млечен път, не се е обособило в звезди веднага след като самата галактика се е уплътнила, тъй като процесът на образуване на звезди продължава и до днес. Някои от звездите, които веждаме по небето, в действителност са се образували първи. Голяма част обаче са се формирали в последствие след първичната галактична кондензация. Сред тази по – голяма част е и Слънцето. Кога са се образували планетите и как е възникнала Слънчевата система, са въпроси, които днес нямат все още напълно определен отговор. При това догадки и хипотези съвсем не липсват.
Малко животни вълнуват човешкото въображение, така както мечките го правят. Мечките стоят изправени на два крака, очите им са отпред на главата, ходят на цялото си стъпало, хващат нещата с пръстите на крайниците си, люлеят и прегръщат малките си на ръце, както ние го правим, хранят се с каквото ние се храним. Тези подобия са забелязани още в ранна човешка история и затова не е чудно, че мечките ни вълнуват така силно. Повечето култури имат или са имали ритуали, асоциирани с мечките. Мечките или техни части са имали специална магическа и медицинска сила. Тези сили не идват само от мечата сила или хитрост, въпреки че скандинавците казват, че мечките имат „ силата на десет мъже и хитростта на дванадесет”, по-скоро тази магическа сила идва от възможността да бъдат между живота и смъртта. През зимата те се отправят в своите бърлоги и изпадат в зимен сън, продължаващ няколко месеца до пролетта, когато се появяват живи и здрави на открито. Разбираемо е мечките да са символ на цикъла на живота и смъртта и на възкресението. Тяхната поява предвещава новия вегетативен период – пролетта. Освен това женските отиват в бърлогата си за зимен сън видимо сами, а се появават с малки мечета през пролетта. Това в редица култури е символ на непорочното зачатие. Много градове, географски местности, семейства, както и символи на властта – монети, марки и административни книжа, са кръстени или имат знак на мечки. В съвремието ни от една страна мечките са символ на дивата природа, а от друга символ на нарушителя на спокойствието на фермери и животновъди, символ на опасност и дори смърт. През последните столетия мечките са подложени на преследване и убиване от човека, дори това е било правителствено стимулирано. Акт довел до силно застрашаване на съществуването на този вид. Мечките гушкат своите малки мечета , а нашите малки деца гушкат пухкави мечета за лека нощ. Въпреки това мечките изглеждат диви и опасни и много хора се страхуват да се разхождат в гора, където живеят истински мечки. За изчезването на видовете
Кватернерът (на латински Quartarius), известен още като четвъртъчен период, е геоложки период, започнал преди 2.588 ± 0.005 млн. години, който продължава до днес. Дели се на плиоцен, плейстоцен и холоцен. През този период са станали над 20 ледникови – междуледникови цикли. Кватернернерните климатични промени са оказали голямо влияние върху биоразнообразието на земята, основано на промените на глобалната температура и нивото на океана, което е променяло хабитатите и екологичните ниши. Например през Плейстоцена /геоложка епоха, започнала преди 1,806 милиона години и завършила преди 11,500 години/ температурната флуктуация с повече от 15 С е ставала в течение на няколко деситилетия, в резултат температурата в глациалния максимум е била с 21 С по-ниска от сегашната. Тези температурни промени са оказвали влияние на нивото на океаните и моретата, тъй като голяма част от водата е влизала в състава на ледниците и големите полярни шапки. Формирането на огромни ледници, като тези на Лорентидите и Кордилерите в Северна Америка, Фено – Скандинавските и Алпийските в Европа, по време на глациалния максимум, води до спадане на морското равнище с 120 м под сегашното ниво. Това спадане на морското равнище и довело до формиране на междуконтинентални мостове или връзки между територии, които в период на затопляне са отделени едно от друго с вода. Такива са Беринговия мост, свързващ Евразия и Северна Америка, моста, свързващ Британските острови и южния край на Скандинавия с Европейския континент, позволили на редица видове да се разселят на нови територии. В отговор на тези глобални климатични промени, много видове са принудени да търсят подходящия за тях хабитат, в резултат през отделните цикли, популациите са разширявали или свивали своя ареал на разпространение. Тези които не са могли да търсят свой нов хабитат, са били принудени да се адаптират или да умрат.
Най-същественото и безпрецедентно събитие е описано за края на последното заледяване. В края на Плейстоцена и началото на Холоцена над 60% от сухоземните гръбначни животни с големина над 44кг измират. За това масово измиране на мегафауната има три основни хипотези – климатичните промени, избиване от човека, вирусна инфекция. Измиране на видове за целия кватернерен период е бил нещо обичайно, тъй като не малко видове не са могли да се адаптират или да мигрират в по-топли географски ширини по време на продължителните заледявания. Необичайното за масовото измиране на мегафауната от края на последния глациален максимум е това ,че са измрели масово видове над 44 кг телесно тегло, което показва, че е се случвало избирателно, още повече, че е засегнало само сухоземните животни. Отворен остава въпроса какво е било по-различно на последното заледяване , спрямо предишните, за да доведе до такова масово измиране на мегафауната. Редица автори смятат, че основната причина е комбиниран ефект от климатичните промени и ловуването от човека, но безпорно е факт , че човешката намеса върху биотата е имала и продължава да има силно негативен ефект.
По време на ледниковите периоди, популациите на видовете са се свивали и се ограничавали на юг, където е имало по- подходящи условия на живот, обособявайки отделни клонове /рефугиуми/ . През периодите на затопляне видовете реколонизират северните територии , като така разширяват чувствително своя ареал на разпространение. Модела на разширяване и свиване на разпространението на видовете е описан като модел за Европа от Hewitt/ и един от моделните видове е кафявата мечка. Филогеографските изследвания на основа на митохондриална ДНК дават възможност да се опише начина на разпространение на видовете по време на глациалните цикли. На схемата е показано филогеографското разпространение на кафявата мечка след последния глациален максимум. По време на заледяването се обособяват три рефугиума – Иберийски, Итало-Балкански и източен евразийски ,който е с неуточнено местоположение. Иберийския и Итало-Балканския са били ситуирани съответно на Пиринейския / Иберийски/ , Апенински и Балканския полуострови. От тях при климатичното затопляне започва разселването на вида в Европа и Азия на север, както е показано.
ЕВОЛЮЦИЯ на МЕЧКИТЕ
Откъде са произлези мечките? Еволюцията на мечките, каквито ги знаем сега, започва преди около 38 млн години. Техен прародител е еволюирал в семейство малки бозайници Миациди (Miacidae). Мечките и канидите са произлезли от тези миациди. Семейството на мечките се отделя от миацидите със своя предшественик най – стария род Ursavus, който имал големина на овчарка. Мечките (Ursidae) формират отделно семейство в разред Хищници Carnivora. Семейството се поделя на три подсемейства: на гиганската панда Ailuropodinae, на очилатата мечка (Tremarctinae) и на същинските мечки (Ursinae). Последното подсемейство включва останалите 6 вида днес живеещи мечки, всички те имат подобни външни белези и сходен начин на живот. Най – стария намерен фосил на полярна мечка е датиран отпреди 130 – 110 хил.год. Фосилите на полярната мечка показват, че преди 10 – 20 хил год кътните зъби на полярните мечки са вече значително променени и различни спрямо тези на кафявата мечка. Счита се, че полярната мечка се отделя и дивергира от популацията на кафявата мечка по време на плейстоценските заледявания. По последни изследвания това вероятно е било преди около 600 хил год, а не както доскоро се е смятало, че обособяването на новия вид е станало преди около 130 хил год.
Фиг. 1 Филогения на сем Ursidae, получена на основа на секвениране на региони от мтДНК на съвременни и фосилни видове. Включено е дивергентното време в млн.год.
Първоначално се отделя гигинтската панда преди около 17,9 – 21,1 млн год. Следващата дивергенция е на подсем на очилатата мечка Tremarctinae, което включва и късомуцунестите мечки , вече изчезнали видове. Тази дивергенция е настъпила преди около 12,4 – 15,6 млн год. За останалите мечки дивергенцията е доста припокриваща се и единствено се откроява дивергенциата на клона на кафявата мечка. В клона на кафявата мечка дивергенцията между пещерната и кафява мечка е била преди около 2,4 – 3,1 млн години. Фиг. 1Времево разпространение на фосилни, изчезнали и съвременни видове мечки. Фосилните и изчезналите видове са означени с кръст. Вертикалния сив правоъгълник дава времето на масовата експанзия на видовете на границата на миоцен – плиоцен.
Преглед на съвременните видове мечки.
ГОЛЯМА ПАНДА, Бамбукова мечка, Giantpanda, Ailuropoda melanoleuca
Гиганската панда е вид мечка, живееща в централната и южна част на западен Китай. Разпространението на вида е ограничен за Сичуан, Саанци и Гансу китайски провинции, в планинските гористи местности, където расте бамбук. Тялото е с характерна мечешка форма, къса опашка и оцветяване на козината в бяло и черно, типично за вида. Въпреки, че спадат към Хищниците, основното меню на тези мечки е бамбука, макар че по изключение могат да се хранят и с плодове, корени, малки птички, гризачи и насекоми. Във връзка с храненето с бамбук, пандата има редица развити приспособления. Китна кост е удължена и действа като палец за захващане на бамбук, стомаха им е силно мускулест, в помощ на стриването на бамбука, зъбната система също е приспособена за хранене с твърдите бамбукови пръчки. Именно зъбната система, черепни особености и начина на хранене са причина дълго време вида да се отнася към Енотови, заедно с малката панда.
Пандата един от най – известните и най-лесно отличимите видове в природата, но и един от най-застрашените от антропогенната дейност. Вследствие на изсичането на горите, вида е ограничен в изключително малка територия и се счита за застрашен от изчезване вид съгласно червената книга на IUCN. Включен е в логото на WWF / World Wildlife Fund/ като символ на природозащита. Китайците виждат в пандите символ на мира и затова полагат изключителни усилия за тяхна защита и запазване на хабитатите.
Пандата живее поединично, като маркира територията си със секрет, отделен от мирисни жлези и драскайки маркери по дърветата. Мъжките големи панди имат почти двойно по-голям ареал от женските, като припокриват територията на няколко женски, на които държат правата за чифтосване. Тъй като бамбука няма висока хранителна стойност, голямата панда трябва да яде повече от 30 кг бамбукови листа, клонки и стъбла всеки ден, като това грубо съставлява 40% от телесното им тегло. По тази причина пандите прекарват между 12 до 15 часа на ден в дъвчене на бамбук, което правят в седящо положение, захванали клонките с предните си лапи. Въпреки че пандите имат вид да прекарват целия си ден в дъвчене на бамбук или спане, те са относително пъргави и могат добре да се катерят по дарветата и дори и да плуват, ако се наложи. Хранят се с над 30 вида бамбук, като използват различни части от растението. Макар бамбука да съставлява 90% от менюто, понякога могат да разнообразят и с треви, плодове, малки птици, гризачи.
Размножителния период е между март и май, когато женската показва на мъжкия , че е готова за чифтосване като започва да издава серия от ревове и звуци, за да го привлече. След периода на бременност , която продължава около 5 месеца, женската голяма панда ражда едно, рядко две малки в основата на кухо дърво или процеп в скалите. Малките на пандата са неразвити, с големина около 15 см и тежат 100г, слепи и голи. Те започват да пълзят едва след 3 месеца. Дори и да роди две малки, майката се грижи за едно, което се захваща за козината и на гърба до шестия месец. Пандата отбива малките след около година, но те могат да продължат да са около нея до 18 месечна възраст. Понякога малките могат да останат при майките си няколко години, докато тя не забременее отново, когато са принудени да напуснат нейната територия и да установят своя.
Пандата е световно известна с оцветената в бяло и черно козина, но има една популация в провинция Саанци, която е в кафяво и бяло. Пандите комуникират помежду си със серия от звуци, като са разпознати около 11 различни звуци. Гиганската панда пигмей A. Microta, сега изчезнал вид, също е бил разпространен за същия ареал, както и съвременната панда
АЗИАТСКА ЧЕРНА МЕЧКА Asian black bear Ursus thibetanus
По старото име на азиатската черна мечка е Selenarctos thibetanus, което означава лунна мечка от Тибет. Фосили от този вид са намирани на територията на Европа, но сега видът е ограничен само в Азия – тясна ивица от югоизточен Иран на изток през Авганистан и Пакистан, през Хималаите в Индия до Мианмар. С изключение на Малайзия, черни мечки се срещат във всички страни на Югоизточна Азия. Отделни популации живеят в южните части на Руския далечен изток , Корея, Япония – острови Хоншу и Шикоку, Тайланд и Хайнан. Предпочитаемия хабитат е листопадни гори, на височина до 3 500 м.
Азиатската черна мечка прилича на външен вид на кафявата мечка, но изглежда малко по – слаба и с по- елегантен силует. Черепа е относително малък, но масивен. Въпреки, че се храни предимно с вегетарианска храна, челюстите не са видоизменени така, както на пандата. Черната азиатска мечка, за разлика от полярната, има по – добре развита горна част на тялото, във връзка с катеренето по дърветата. В сравнение с кафявата и американската мечка, краката са по-слаби и къси. Козината е черна /рядко кафява/ , с ясно видим V – образно бяло петно, оприличавано на лунен полумесец. Около врата имат яка от по – дълга козина.Имат много добре развито обоняние, но зрението и слуха са слаби.
Като останалите видове мечки и азиатската черна мечка живее поединично и се събира с други единствено за размножаване или когато се съревновава за желани територии. Въпреки по-малките си нокти, тези мечки са добри катерачи и прекарват по-голяма част на дървета, където си правят груби «гнезда» от пречупени клони, берат плодове и ловят малки животни. В по-студеното време в по-северните региони, азиатските черни мечки също изпадат в зимен сън и също есента се хранят със жълъди, орехи и други ядки, даващи повече мазнини, с цел да се оформи запасен подкожен слой за зимния сън. Изпадат в зимен сън от ноември до април, а в някой части на Русия от октомври до май.
Навършват полова зрялост около 4 – 5 година и се чифтосват през топлите летни месеци юни – юли. След период на бременност дълаг от 6 до 8 месеца, през март април се раждат от 1 до 4 малки / най-често 2/ в топлата и защитена бърлога, осигурена от женската. В зависимост от региона тези бърлоги могат да бъдат открити в близост до реки или скални цепнатини. Тези видове, живеещи в по-топлите региони, които не изпадат в зимен сън , също правят бърлоги, където да родят и отгледат малките си. Малките се раждат голи, слепи и безпомощни. Майките ги кърмят до 6 месеца и след това започват да се хранят с твърда храна. Въпреки това някой малки остават до майките си до 3 годишна възраст. В плен Азиатската черна мечка живее до 30 години, в природата не са срещани по-възрастни от 25 години.
Въпреки, че са класифицирани като хищници, като всички мечки и азиатската черна е всеядна, хранейки се с растителна храна и малки животни. Прекарва по- голяма част от времето си в хранене с ядки, семена, плодове, треви, листа, билки, малки насекоми – мравки и термити, рядко птички и грицачи, като всички те съставляват основното и меню.
Азиатската черна мечка е известна и други свои популярни в Азия имена, като бялото петно на гърдите с формата на полумесец и дава името Лунна мечка. Тези мечки са активни основно през ноща, като през деня спят по дървета в груби гнезда от клони или в големи хралупи на кухи дървета, както и в скални процепи, излизайки навън през ноща. По последни изследвания в менюто им влизат повече от 160 видове плодове и ядки.
Има исторически данни , че човека е преследвал и убивал тези животни от повече от 3000 години насам, главно заради лапите и жлъчката, която изсушена се е използвала като лекарство в китайската медицина, като се е вярвало, че има редица лечебни качества. Отглеждани са в специални ферми за жлъчки в Китай и Виетнам. Тази индустрия е довела до драстично намаляване на популациите и заедно със загуба на хабитата, вследствие на изсичането на горите водят до сегашното им състояние на застрашен от изчезване вид. Макар и рядко тези мечки са нападали фатално хора и това също е водело до тяхното изтребване. В Пакистан са отглеждани като домашни любимци за борба с кучета, наричана мечо хапане. Преди да бъдат пускани в битка, на мечките са вадени зъбите и ноктите и така те не са можели дори да се защитят срещу нападението на бултериерите.
Според Червената книга на IUCN , тази мечка е застрашена от изчезване. Само за последните 30 години, популациите са намалени с 49%. Според последни данни в природата съществуват не повече от 50 000 мечки. Въпреки наложените забрани, продължава тяхното избиване или разрушаване на хабитатите, мизерното им съществувание в мизерните ферми за жлъчка или за кучешкото мечо хапане.
Малък подвид, тежащ между 60 – 120 кг за възрастен мъжки и 40 – 100 кг за възрастна женска. Дължината на тялото варира от 110 – 140 см. Нямат плътната козина около врата, характерна за другите подвидове и има по-тъмен цвят на муцуната.
ГАЛЕРИЯ:
АМЕРИКАНСКА ЧЕРНА МЕЧКА – Americanblackbear – Ursus americanus
Американската черна мечка е най-малката от трите вида мечки, обитаващи територията на Северна Америка. Тази мечка се среща само в Северна Америка, обитаваща различни горски хабитати. Тя е не само най-широко разпространения вид за Северна Америка, но и най- многобройния. Американската черна мечка е един от двата вида които, за разлика от останалите видове мечки, не са описани като застрашени или изчезващи от червената книга на IUCN. Определени са 16 подвида на този вид, които основно варират по големина и цвета на козината.
Въпреки името си, цвета на козината на тези мечки варира от черен, тъмно червеникав или кафяв, с различни просветлени нюанси, в зависимост от района на местообитание.
Имат къса, гъста козина, в сравнение с кафявата мечка по-малки очи, по-големи уши, нямат гърбица на рамената, по-къси крака и нокти, вследствие на което са по-добри катерачи и по-подвижни. Имат много добро развито обоняние, слаби слух и зрение.
Въпреки, че сега видът е широко разпространен в Северна Америка, в миналото ареала му е обхващал целия континент, на всеки подходящ горски хабитат. Сега видът е ограничен от северна Канада, през западен и части от източен САЩ, надолу през северно Мексико. Предпочитат топли и сухи обрасли с храсти гори в Мексико, влажни и блатисти гори юго източен САЩ, иглолистни гори в Канада, дори и нискорастителната тундра на север.
Черната мечка живее поединично, активна нощем. Прекарва по-голямата част от времето си в търсене на растителна храна, ядки, семена, които яде в голямо количество, с цел да натрупа подкожни мазнини за зимата. Когато застудее и настъпи зимния период те се отправят в своята бърлога, цепнатина в скалите, кухо дърво, където прекарват в полусънно състояние. Въпреки,че това не е същински зимен сън, и въпреки, че температурата на тялото им се понижава, са способни да прекъснат дрямката си и да потърсят нещо за похапване.
Мъжките и женските черни мечки се събират за кратко през пролетта, след което се разделят и започват активна подготовка за зимния период. След 7 месечна бременност, женската ражда 1 до 5 малки мечета. Това обикновенно става в края на зимата. Малките са слепи, голи и безпомощни и се закрепват за майката, за да се топлят. Мечетата остават близо до майките си до 2 годишна възраст, като се обучават в умения да преживяват сами.
Въпреки, че са класифицирани като хищници, черните мечки имат предимно вегетарианско меню. Адките, семената, коренищата, тревите, листа, клонки, съставляват над 80% от основното им меню, което слабо се разнообразява с малки животни – насекоми, гризачи. Рядко са известни случай да преследват сърни или да ловят риба, главно в тези региони , където няма мечки гризли. Поради голямото си тегло, мечките прекарват живота си основно в хранене, за да съберат достатъчно резерви за зимата.
Черните мечки като цяло нямат много врагове, поради големите си размери. Малките често стават жертва на лисици, хищни птици и дори други черни мечки. Въпреки ,че между човека и черните мечки са установени относително миролюбиви отношения, има случай на преследване от фермери, страхуващи се за животните , които отглеждат. Рядко има случай на намаляване на популации, вследствие унищожаване на хабитатите. Като цяло видът е свикнал да живее в близост до населени места и е дружелюбен. Известен е с това ,че може да влезе в дома или в колата, в търсене на храна. Чести са за националните паркове и не са малко случаите на вземане на храна от запасите на къмпингуващите там.
Подвидове Американска черна мечка
Sub-species Име
Разпространено име
Разпространение
Описание
Ursus americanus altifrontalis
Олимпийска черна мечка
Северозападното крайбрежие на Тихия океан от централна Британска Колумбия през северна Калифорния и прилежащата суша до върха на северно Айдахо, Британска Колумбия
Ursus americanus amblyceps
Новомексиканска черна мечка
Колорадо, Ню Мексико, западен Тексас, източната половина на Аризона и северно Мексико, югоизточна Юта
Ursus americanus americanus
Източна черна мечка
Източна Монтана до Атлантическия бряг, от Аляска на юг и изток през Канада до Атлантическия океан и южно до Тексас
За някой региони се смята, че популацията му се увеличава, един от най-големите подвидове, всички са с черна окраска на козината, Обикновен за Източна Канада и САЩ. Много рядко може да се видят индивиди с бяло петно на гърдите
Ursus americanus californiensis
Калифорнийска черна мечка
Планините на южна Калифорния, северно през Сентръл Вали до южнен Орегон
По-голяма от своите континентални сродници,с размери вариращи от 90 до 270 кг. Имат един от най-дългите периоди на зимен сън от всички северноамерикански мечки
Приблизително 10% от популацията на този подвид е с бяло или кремаво оцветена козина, дължащо се на рецесивни алели и затова се наричат мечки Дух или Кермоде. Останалите 90% са с нормална черна окраска
Ursus americanus luteolus
Луизианска черна мечка
Източен Тексас, Луизиана, южно Мисисипи
Има относително дълъг, слаб и плосък череп, пропорционално големи кътници
Ursus americanus machetes
Западномексиканска черна мечка
Северно – централно мексико
Ursus americanus perniger
Кенайска черна мечка
Полуостров Кенай, Аляска
Ursus americanus pugnax
Далска черна мечка
Остров Дал , Архипелаг Александър , Аляска
Ursus americanus vancouveri
Ванкувърска островна черна мечка
Остров Ванкувър , Британска Колумбия
Какво прави бялата мечка бяла?
При подвида мечка Кермоде или мечката дух Ursus americanus kermodei , приблизително 10% от популацията на този подвид е с бяло или кремаво оцветена козина. Учените са направили изследвания на ДНК от косми на бели и черни индивиди на този подвид, за да установят каква е причината. Установяват една точкова мутация на 1 нуклеотидна двойка в гена на меланокортин 1 рецептора, който всъщност обуславя какъв цвят козина да се произвежда от клетките. Тази мутация променя белтъчната структура на рецептора, което стопира производството на пигмента и оставя мечките бели.
Тази неголяма мечка е уникална сред своите родственици, поради основното им меню насекоми – мравки, термити и др. Бърнестата мечка обитава горските и тревисти райони на Индийския полуостров. Има дълга, пухеста козина, с удължени около рамената участъци, оцветена в черно. Рядко се срещат в канелен, червеникав цвят на козината. Отпред на гърдите имат бяло или кремаво петно в U’ или Y’ форма. Характерна е източната муцуна в по-блед цвят. Формата на муцуната, голите бърни и липсата на горни резци са адаптация за хранене с насекоми. Предните лапи са извити навътре и имат дълги, леко закривени нокти, предназначени за ровене и копаене.
Живеят поединично, като маркират територията си, драскайки по дърветата. Активни са и през деня и през ноща, с изключение на майките с малките, които се придържат към дневна активност. Времето за чифтосване е от юни до юли. След бременност, продължаваща 6 – 7 месеца, се раждат обикновенно 2 малки. Майката стои до малките до третия им месец, когато те са малко по-големи и се движат навън с нея, захванати за гърба и. Малките остават при майка си често повече от 2,5 до 3 години, и затова женските раждат на интервали не по-малки от 3 години.
Бърнестите мечки са уникални със своето меню, в което влизат основно насекоми – най – вече термити и мравки. Разриват с предните си дълги нокти термитника или мравуняка и завират дългата си муцуна вътре, като първо издухват пръста и праха, преди да започнат да засмукват мравките. Хранят се също и с мед, изтърпявайки ужилванията на пчелите, докато вземат пчелните пити. Менюто си разнообразяват и с яйца, мърша, растителност и плодове.
Основните заплахи за този вид са загубата на местообитание и бракониерството. Местообитанията се разрушават при изсичането на горите, заради насаждането на монокултурни плантации, селища на бежанци, строежи на селища, увеличаване на земеделските земи, пътища. Бракониерството е главно от комерсиална гледна точка, мъжките репродуктивни органи се считат за афродизиак, костите, зъбите, ноктите се използват да пазят от зли духове, мечата мазнина се използва за лек от народната медицина. Малки мечки се пленяват за дресиране на „ танцуващи мечки” и това остава основна заплаха за някой популации. И звестни са отделни случаи на нападение на хора от мечки, довело до тяхна смърт. Единствения естествен враг е тигъра Panthera tigris и леопарда P. Pardus. Бърнестата мечка е регистрирана в 174 защитени ареала в Индия, които включват 46 Национални паркове и 128 Защитени зони. В червената книга на IUCN е включена като уязвим вид.
Шри Ланските бърнести мечки имат по – къса козина, което ги прави по-малко пухкави. Имат и по-малки размери на тялото и зъбите. Срещат се и видове без бялото петно на гърдите. Поради мащабното изсичане на горите и превръщането на им в плантации за кафе и чай, популацията на бърнестата мечка в Шри ланка значително намалява . Сега са ограничени в северните и източните низини.
В Индия са разпространени на отселни популации, предпочитаемо в горски хабитати. Не се срещат по високите планини на Химачал Прадеш, Джаму и Кашмир, както и в северозападните пустини на Раджастан и простроните полета на юг. Тази мечка сега е най-разпространената в Индия, срещайки се в Сивалиск / също в Пакистан/, ниските хълмове граничещи с Хималаите, от Пинджаб до Арунчал. Изолирани са от популациите на Непал, заради обработваеми земи. Бърнестите мечки в Непал са ограничени главно в Терай, южна ивица гори и поляни, граничещи с Индия. Няколко изолирани популации се счита, че все още съществуват в Читагонг и Силхет, региони на източен Бангладеш.
Известна също като медeна мечка, заради любовта си към меда, слънчевата мечка е най-малката , най-малко позната и най-рядката мечка.
Слънчевата мечка е малка мечка, обитаваща гъстите тропични гори на Югоизточна Азия. Тя е най-малката мечка в света и също така уникална с това, че е адаптирана за живот в джунглата и прекарва живота си основно по дърветата. Известна е и като Малайска слънчева мечка, медена мечка, има редица отличителни белези, най – забележителния е оранжевото U образно петно на гърдите, откъдето и е името на тази мечка. Това са редки и почти неуловими животни и практически малко се знае за тях и начина им на живот, освен че драстично намаляват популацията си , заради лова и разрушаването на хабитатите.
Освен най- малки, те са и с най-късата козина. Цвета и варира от черен, тъмно кафяв или сив, а петното на гърдите е бяло, жълто или оранжево, като формата му е различна – полумесец, кръг или неправилно по форма. Муцуната е по-светла, къса, с къса козина и малки уши. Имат дълги закривени нокти, които им помагат да се катерят по дърветата и да разриват термитници. Един от най-отличителните белези е техния дълъг повече от 25 см език, с който лесно загребват ларви или мед.
Слънчевите мечки се срещат в няколко азиатски страни, но поради човешката дейност, са ограничени в определени хабитати. Срещат се на островите Борнео и Суматра, в горите на Малайзия, Тайланд, Камбоджа, Лаос и Виетнам, като има и няколко откъснати популации в южен Китай. Предпочитат нискоразположени дъждовни гори, където намират разнообразна храна и успешно се прикриват от врагове. Поради обезлесяването тяхното разпространение е драстично намалено.
Основна част от живота си прекарват по дърветата, като дори спят в големи, груби гнезда, направени от пречупени обрасли клони. Катерят се сравнително бързо с помощта на закривените си нокти. За разлика от повечето си сродници, тези мечки са дневни животни и са активни през деня, който прекарват в търсене на храна, а си почиват през нощта високо в техните гнезда на дърветата.. Изключение правят само тези мечки, които са силно афектирани от човешката дейност и са принудени да бъдат активни през нощта. Живеят поединично и се събират само за да се чифтосват.
Подобно на други животни, живеещи в тропиците и слънчевите мечки са способни да се чифтосват и раждат целогодишно.Бременноста продължава от 3 до 8 месеца, варираща индивидуално и от наличието на храна. Раждат се до 3 малки , обикновенно едно в гнездо, което женската прави високо на някое дърво или в голяма кухина. Женската се грижи за малките и ги защитава, учи ги на умения да оцеляват сами в джунглата, когато пораснат и станат самостоятелни. Малките остават при майка си до около 2 години, след което установяват самостоятелна територия. Женските сързяват на 3 години, а мъжките на 4 години. Продължителността на живота им е около 25 години.
Слънчевата мечка има много разпространени имена едно от които е мечка – куче.
Поради това, че обитава тропиците тази мечка не изпада в зимен сън.
Менюто им включва основно плодове, семена, ядки, растения, насекоми – термити и мравки, яйца, мед, малки животни като птички и гризачи и то тогава, когато растителната храна е недостатъчна. Като другите видове мечки кътниците им са по – плоски, поради хранене не с месо, а с растителна храна.
В природата основен техен враг е тигъра. Малките често биват нападани от змии, големи хищни птици.Най-големия техен враг остава човека с неговата дейност по обезлесяване, което води до загуба на хабитата им. Живеещите до земеделски насаждения с африкански маслени палми мечки обичат да навлизат в плантациите и да ядат младите филизи, с което нанасят вреда на плантациите. Затова често мечките са преследвани от фермерите, които се страхуват за своите посеви.
Преди години тези мечки са били обект на лов от хората, главно заради части на тялото и най-вече жлъчката, използвани от местната медицина. Поради малкия им размер и хрисимия характер тези мечки са били улавяни и продаване като атрактивни домашни любимци в различни части на света.
Сега Слънчевата мечка е включена в Червената книга на IUCN като уязвим вид. Смята се , че популацията на тези мечки е намаляла с повече от 30% през последните няколко десетилетия. По последни данни, макар да се считат за не особено точни , в природата живеят не повече то 1000 индивида. С цел тяхното опазване, правителството е забранило нелегалната сеч, отстрела и бракониерството в районите където се знае , че съществуват.
ОЧИЛАТА МЕЧКА SpectacledbearTremarctosornatus/други имена – укуко, юкумари, укумари/
Очилатата мечка е позната и като Андска мечка, основно защото е характерна за Андите в Южна Америка. Територията и се простира от северозападна Аржентина, Перу, Венецуела, Колумбия, Еквадор, източна Боливия.
Храни се основно с плодове и филизи, които събира от дърветата или от земята. Менюто включва допълнително мед, насекоми, малки бозайници, влечуги и рядко добитък. От културните растения предпочитат захарна тръстика, царевица.
Очилатата мечка е иднствения оцелял до днешно време вид от сем. Tremarctinae, като за една от причините за неговото оцеляване се счита възможността му да се катери по дърветата и приспособимостта му към разнообразни хабитати от сухи тревисти поляни до влажни високопланински гори. Към това семейство са спадали: видът Plionarctos, който е живял в Северна Америка през късен миоцен и ранния плиоцен, видът Tremarctos floridanus описан за Северна Америка за периода на Плиоцена – Плейстоцена, късомуцунестите мечки от периода на Плиоцена – Плейстоцена, Arctodos simus Гигантската късомуцунеста мечка и Arctodus pristinus Малката късомуцунеста мечка. Arctodos simus Гигантската късомуцунеста мечка се счита за най-голямата живяля някога мечка. Тя е била по-голяма от съвременната мечка гризли.. Ако се съди по намерените скелети днес, височината на арктодуса е от 1,5 до 1,8 m, а изправена достига 3,5 m. Тежала 600 kg, а най-големите екземпляри достигали 1136 kg. Имала е силно изразен полов диформизъм
Живеят предпочитаемо в дъждовните гори и джунглите на Андите, избирайки по – влажни и по залесени ареали. Но при търсене на храна са срещани от слабо зелесени сухи низини до високопланински гори. Активна е денем, за разлика от повечето видове мечки, които са активни нощем. Счита се, че тази мечка няма своя територия, но пази дистанция от по – големи животни и особено от човека. Не е агресивна , обикновенно е хрисима и приятелски настроена при срещите с хора. Все пак женските са по-агресивни, когато защитават малките си и макар този вида да е със среден ръст, възрастните достигат 1, 5 м в изправено положение и използвайки мечата си сила биха могли да бъдат опасни, когато защитават своите малки. Козината е черна, рядко тъмно кафява или червеникаво кафява окраска с характерно светло бежово петно около очите , врата и гърдите, откъдето им идва и името. Мъжките са почти два пъти по-големи от женските . Тежат от 100 до 200 кг, докато женските от 35 до 82 кг. . Размножоват се целогодишно , но пика е от април до юни. Мъжките и женските се събират за седмица – две, след период на бременност, продължаващ от 5,5 до 8,5 месеца се раждат от едно до три малки, най-често две. Малките се раждат слепи и безпомощни, тежащи до около 300г. Майката се грижи за тях до едногодишна възраст. Тези мечки не изпадат в зимен сън.
Популацията на очилатата мечка е намаляла числеността си през последните деситилетия. Основните причини от една страна са защото фермерите смятат, че тези мечки нападат добитъка им и от друга поради унищожаване на хабитатите. Жлъчката на тези мечки също се счита ,че има лечебно действие и често са убивани и по тази причина. Сега Очилатат мечка е включена в Червената книга на IUCN като уязвим вид
ПОЛЯРНА МЕЧКА, Бяла мечка, Морска мечка, PolarbearUrsusmaritimus/други имена: нанук на инуит, юпук в Сибир, ошкуй в Коми, исбьорн – ледена мечка в Свалбард/
Полярната мечка е голяма мечка която обитава ледените полета в Арктическия океан. Тя е най-голямата съвременна мечка. /изключение прави Кодиак кафява мечка в Аляска, която достига подобни размери/, като мъжки достига тежина около 700 кг. Тази мечка е най-близкия родственик на кафявата мечка е наречена морска мечка, понеже е срещана най-често по крайбрежията, в близост до водата, освен това тя е много добър плувец, виждана навътре в океана на повече от 100 мили от най-близката суша.
Константин Джон Фипс, втори барон Молгрейв, е първия описал този вид мечка през 1774 г, избирайки и името морска мечка поради условия на живот ,които е видял да води. Разпространението на този вид е най-много повлияно от глобалното затопляне и затова полярната мечка е станала символ на климатичните промени. Намаляването на популацията е способствано също от човешката дейност – лов, замърсяване, сондажна дейност за добив на нефт и др.
Полярната мечка достига 2 метра на дължина и тежина поне половин тон. Женските са по-малки дори до два пъти. Те са от малкото големи бозайници, адапитрани за живот при неблагоприятните арктически условия. Във връзка с живота на леда са развили редица приспособления – дебела козина с много пух и по дълги косми, които предават слънчевата топлина на черната кожа, която абсорбира топлината, дебел подкожен слой мазнина. Едро, здраво тяло с големи предни лапи, с които чудесно плува. Козината достига дори между пръстите, за да топли. При ходене върху леда ,лапите им имат чудесно сцепление върху него. Шията им е по- дълга, за да остава над водата като плуват. Муцуните са издължени, а ушите малки. Въпреки, че са едри могат да развиват до 40 км/ч скорост на леда и до 10 км/ч плувайки.
Белите мечки са срещат циркумполярно, като достигат на юг до Хъдсън Бей и Ню Фаунленд. Около 60% от тях се срещат в северна Канада, а останалата част обитават Гренландия, Аляска, Свалбард, Русия, кадето се срещат недалеч от брега, изминаващи дълги разстояния по леда. Живеят поединично и са доста добри ловци както на сушата, така и във водата. Могат да се гмуркат, за да хванат плячката, като държат очите си отворени и задържат дъха си до 2 мин. На сушата или преследват плячката или стоят с часове до някой дупка в леда, дакато не хванат плячката, която се е подала да подиша над леда. Най- любимата и високоенергийна храна за тях са тюлените. По време на летния период мигрират далеч на север по леда, за да търсят тюлени. Белите мечки са едни от най-едрите хищници на земята ,които редовно трябва да си набавят храна, за да поддържат енергийния си баланс и дебелия подкожен слой мазнина. Кожата и маста на тюлените са една от любимата им храна, като оставят месото за други по-малки полярни животни като полярната лисица. В менюто на мечките още се включват птици, риба, плодове, през лятото понякога северни елени. Труповете на големи животни като моржове, китове, тюлени също могат да се окажат важен източник на храна. Миризмата на трупа се усеща от много далечни разстояния от белите мечки.
Размножителния период е април до май, след бременност, която варира по продължителност в зависимост от здравето на женската и наличието на храна, след около 9 месеца се раждат от 1 до 4 малки, в бърлога, която тя изрива в снега или почвата. Малките са слепи, голи и безпомощни, тежат не повече от половин килограм. Женските влизат в бърлогите си късна есен и не се показват с малките докато не настъпи пролетта. Малките остават с майка си до 2 – 3 годишна възраст като се обучават да ловуват и да се борят, подготвят се за самостоятелния живот като възрастни. Продължителност на живот около 25 години, като е регистрирана в плен до 43 години
Поради големината си нямат естествени врагове, освен сблъсъка с други мечки от техния вид в защита на малките. Най-големия враг на полярните мечки е човека, който активно ги е ловувал по данни от 1600 до 1970 год, когато се приема международния закон за забрана на лова на този вид. Освен ловя, други негативни фактори са глобалното затопляне, замърсяването и нефтения сондаж. Някой експерти твърдят, че ако се задържат тези условия и поради ниската, по-скоро бавната репродуктивност на белите мечки, те ще изчезнат напълно след около 30 години. По последни данни популацията на белите мечки наброява 20 000 – 25 000 индивида.
Включена е в Червената книга на IUCN като уязвим вид
Тази година, 2013 , е обявена от природозащитници за година на полярната мечка.
Картата показва разпространението на 19 субпопулации полярна мечка.
За кореннота население на Арктика, белите мечки имат изключително културно значение. Открити са останки от лов отпреди 2500-3000 години, както и пещерни рисунки отпреди 1500 години в Чукотския п-в. Предполага се, че уменията на тези народи в улова на тюлени и строежа на иглута е заимстван от полярните мечки. Има много инуитски и ескимоски легенди и предания за тези мечки. За съзвездие полярна мечка заобиколена от кучета, които слезли долу на земята. Многото общи подобия между мечките и хората са вероятната причини да се смята, че духовете на хората и мечките са взаимозаменяеми. Всички легенди показват дълбокото уважение към тези животни, неразривно свързани с живота на полярните хора.
Наричана на староанглийски още бруун или бруин, което също означава кафява, в Дивия Запад, мечките гризли ги наричали Олд Ефрейм /“Old Ephraim“ / или Мокасиновия Джо /“Moccasin Joe“/, латинското наименование идва от латинската дума мечка – ursus и старогръцката дума мечка – arctos. Кафявите мечки са широко разпространени в голяма част от Северна Америка и Евразия в гористи и планински региони. Този вид е национално животно на много щати в Северна Америка и страни в Европа и Азия.
Кафявата мечка е адна от най – големия вид мечки. Достигат 2 – 3 метра дължина и изглеждат огромни, когато се изправят на задните си лапи. На тежест достигат до 635 кг, като подвида мечката Кодиак, заедно с полярната мечка са най – големия вид мечки.
Имат гъста, платна, дълга козина, която варира в различните региони по оцветяване от тъмно до светло кафява, червеникаво кафява със сребристи краища. Ноктите са много дълги, закривени, предимно тъмно оцветени. Размерите на тези мечки силно варират спряма географскя регион, където живеят, пола, възраста, сезона, и дори имат и индивидуални различия. Този вид е най – вариабилния по окраска и размери от всички мечки. При всички подвидове, мъжкия е по – голям от женската.
Кафявата мечка е нощно животно, но понякога може да бъде наблюдавана рано сутрин и късно вечер. Живее поединично, маркирайки голяма територия, но понякога се събират по няколко индивида, ако е налично голямо количество храна, събират се също, за да се чифтосват. За разлика от други видове, кафявите мечки могат да живеят в една просторна територия по няколко индивида, които едниствено изпадат в кнофликт, за да се съревновават за женската.Мъжките често са агресивни спрямо малките мечета и често ги убиват. В своята среда, кафявата мечка е доминиращ хищник и единствено човека е заплаха за нея. Има големи рамена, със силни мускули, което я отличава от другите видове мечки. Силните раменни мускули обуславят силен удар на предните лапи и то такъв, че е способен да пречупи кост, което в добавка със силните, остри, дълги до 15 см, закривени нокти е причина за легендите на мечата сила и страшния мечи замах.
Менюто им е изключително разнообразно, в зависимост от обитаемия регион и наличието на храна. Естествено, че най – достижимата храна е най – предпочитана. Въпреки, че се считат за опасни хищници, кафявите мечки предпочитат вегетарианска храна, риба и малки животни, по – рядко преследват по – големи бозайници като елени. Предпочитаемо се хранят с плодове, семена, ядки, жълъди, шишарки, гъби, млади филизи, корени и други растителни части, мед. С дългите закривени нокти успешно разриват земята, за да достигнат до желаната храна. През есента се хранят усилено, за да се подготвят за дългия зимен сън, който продължава до пролетта.
Размножителния период от средата на май до юни. След оплождането яйцеклетката не се имплантира веднага, а едва по време на зимния сън. Това се нарича забавена имплантация. На пролет женската се появява извън бърлогата си с едно до четири мечета, най – често две. Мечетата се раждат голи, слепи и много малки в бърлогата, докато майката спи. Ако мечката не е натрупала достатъчно подкожна мазнина за зимата и е слаба е възможно яйцеклетката да не се имплантира в матката, а да се резорбира. Малките бозаят до пролетта когато напускат бърлогата и вече тежат от 6 – 9 кг и са достатъчно развити, за да могат да се хранят самостоятелно с твърда храна. Мечетата остават до майките си от 2 рядко до 4 години, през което време се учат да си намират храна и да се защитават. Често малките са прогонени или убивани от мъжкия , който е дошъл за чифтосване или е бил наблизо в тяхната територия. Продължителността на живота на тези мечки в природата е до 28 години. В плен са достигали 48 години.
Отношенията човек – мечка са хилядолетни. По принцип мечката не е голяма заплаха за човека, с изключение когато женската защитава малките мечета. Въпреки това мечките навлизат в селищата на хората в търсене на храна, нанасят вреди на културните посеви и на добитъка. Не са малки и случаите на нападения на хора от мечки по една или друга причина. Човека си остава най – сериозната заплаха за мечките. Вследствие на обезлесяването, замърсяването, лова и бракониерството, числеността на кафявата мечка е драстично намалена през последните няколко века. В момента се счита, че общо кафявите мечки наброяват около 200 000 индивида.Поради тази причина е приета забрана за лова на тези животни, с цел възстановяване на популациите им. В червената книга на IUCN видът е включен като слабо засегнат.
Подвидове кафява мечка
Редица са разногласията по отношение на класификацията на вида. Някои автори посочват около 90 подвида, докато съгласно ДНК анализ различията се свеждат до 5 клона. Скорошни ДНК изследвания показват, че Евразийската кафява мечка и Северно Американската генетично са хомогенни и тяхната филогеография не отговаря на традиционната таксономия. От 2005 са общоприети 16 подвида, дадени в таблицата по-долу.
Subspecies Име
Снимка
Разпространение
Описание
Ursus arctos arctos –Eurasian brown bear
Евразийска кафява мечка
Европа, Кавказ, Сибир и Многолия
Предимно тъмно оцветена козина, рядко с по-светла, средна по размер, тъмни нокти, сибирската е по-голяма от тези живеещи в евразийската част, те се и ловуват по-малко. Среща се в Европа, предимно горски обитател
Ursus arctos alascensis– Alaska brown bear
Аляска кафява мечка
Аляска
Ursus arctos beringianus –Kamchatka brown bear (or Far Eastern brown bear)
Камчатска кафява мечка или Далекоизточна кафява мечка
Камчатски полуостров и остров Парамушир
Много голяма, тъмнооцветена, с тъмни нокти, счита се за предшественик на of U. a. middendorffi.
Западна Канада, Аляска, северозападни САЩ, исторически са съществували и в прериите Грейт Плейнс
Средно до тъмно кафяво, с сиви връхчета на козината. Крайбрежните популации са по- едри – възрастен мъжки гризли тежи средно 364 кг, докато мечка ( женска) от Юкон може да тежи 80 кг. Крайбрежните мечки могат да достигнат двойно теглото на планинските гризли. Високо адаптивни, могат да живеят в плaнинските иглолистни гори, умерени дъждовни гори, сухи тревисти поля и прерии
Ursus arctos isabellinus –Himalayan brown bear
Хималайска кафява мечка
Непал, Пакистан, Северна Индия
Има червеникаво кафява или пясъчна окраска, тази мечка е по- малка от много други кафяви мечки , които могат да се срещнат в Азиатския контитент.
Ursus arctos lasiotus – Ussuri brown bear (or Amur brown bear, black grizzly or horse bear)
Кафява мечка усури, Амурска кафява мечка, черна гризли или конска мечка
Русия – Южни Курилски острови, остров Сахалин, Приморски край, Усури / река Амур региона на юг от Становой хребетRussia: Китай – Хейлонджанг, Япония, Хокайдо
Тази мечка се счита за предшественик на U. a. horribilis
Северно Мексико – Чихуахуа, Коахуила и Сонора, югозападни САЩ – южните части на Тексас, Аризона и Ню Мексико
Счита се за изчезнала, поради масовото отглеждане на добитък в ранчота в САЩ и Мексико. Понеже е доста адаптивен подвид при сухи условия, все още има надежда, че може да са оцелели индивиди, които да живеят или в иглолистнити гори на Мексико или в каньоните на пустинята Сонора
Ursus arctos pruinosus –Tibetan blue bear
Тибетска синя мечка
Тибетското плато
Средна по размер с дълга пухкава козина, тъмна или по – светла, като преходните варианти са най-распространени Козината около врата е по – светла и формира яка. Черепа се отличава с относително плоски хоани, мъдреците са набраздени, груби, големи зъби
Ursus arctos sitkensis
Остров Баранов
Счита се за най – близката до полярната мечка.
Ursus arctos stikeenensis
Британска Колумбия
Ursus arctos syriacus –Syrian brown bear
Сирийска кафява мечка
Кавказ, Сирия, Ирак, Турция / полуостров Мала Азия/, Иран, Афганистан, Пакистан, западни Хималаи, Памир- Алтай, планината Тиен Шан, преди и в Израел
Светло оцветена козина, светли нокти, средна на ръст.
Ursus arctos marsicanus –Marsican brown bear
Марсиканска кафява мечка
Марсика в централна Италия
Останали са около 30 – 40 такива индивида в района на Марсика.
Филогеография на кафявата мечка
Филогеографията е термин въведен през 1987 / Avise et al. ,1987/ и включва проследяването на географското разпространение на генеалогични линии, обикновенно на интреспецифично ниво , използвайки генетичен маркер, най – често последователности от митохондриална ДНК. Митохондриалната ДНК е особено подходяща за такива изследвания, поради относително високата степен на мутации и поради независимото предаване /нерекомбиниращо/ само по майчина линия. По тази причина маркери от мтДНК са широко използвани през последните години за изследване на разпространението на видовете по време на глациалните плейстоценски цикли. От друга страна кафявата мечка е чудесен модел за проследяване на разпространението на бозайниците по време след плейстоценските заледявания. По настоящем вида е широко разпространен в Холарктика и освен това има относително достатъчно намерени фосили отпреди 100 – 150 хил години, които могат да изяснят значително процесите на разпространение.Важен аспект на филогеографията е определянето на монофилетични групи индивиди, които имат общ предшественик помежду си, в сравнение с друга група индивиди – клонове. На основа на последователност от мт ДНК, кафявата мечка е разделяна на няколко генетично различни линии / клона/. Според едни от първите изследвания, съвременните кафяви мечки се разделят на две основни линии – западен клон 1 и източен клон 3а. / Taberlet et Bouvet, 1994/. Западния клон 1 е разпространен в Европа от Испания на север до източна Скандинавия и на изток Италия и Балканския п-в, Източния клон 3а – Русия, северна Скандинавия, източна Европа. Двете имат контактна зона в Скандинавия. Този досегашен модел се счита , че точно описва разпространението на мечките при затоплянето след последното заледяване от три рефугиума – а именно Иберийски, Итало – Балкански и източен / неуточнен/ на север. Досегашния модел предполага, че първи са започнали това разпространение мечките от Иберия, докато тези от Балканите последни, поради по бавното освобождаване от ледниците на Алпите. По – нови изследвания, включващи още по – голяма извадка от проби и най – вече изолирани ДНК проби от фосили, детайлизира и уточнява процеса на постглациално разпространение. На основа на тези изследвания кафявите мечки се делят на 6 клона / номерирани от 1 до 6/ и две именувани по географския регион /Иран и Северна Африка/.
Фиг. 1
а/ Филогеографско разпространение на кафявата мечка по клонове, съответно в сегашно време – по-тъмно оцветената част и допълнително в историческо време по – светло оцветената част.
б/ цветно представяне на хабитатите в Северното полукълбо по време на последния глациален максимум, съответно: вода; лед; тундра/ полярна пустиня; умерен/тропическа пустиня; гори; тревисти полета /прерии/
с/ Вероятни рефугиуми по време на последния глациален максимум и вероятни пост – глациални пътища на разпространение по време на последващото затопляне на климата. : IB – Иберия; IT – Итало – Балкански полуострови; CM – Карпатски планини; CA – Кавказ; UR – Урал; CS – Централен Сибир; NAF – Северна Африка; ME – Среден Изток; SA – Южна Азия; JA – Япония; BE – Берингия; PC – Тихоокеански крайбрежни острови; NA – Северна Америка.
Три генетично различни клона са описани за Северна Америка – 2, 3 и 4. Клонове 2 и 3 се разделят на подклонове 2а, 2в, 3а и 3в. Клонове 2с и 3с са вече изчезнали. Подклоновете са разпространени географски съответно: 3а – северозападна Аляска; 3в – източна Аляска и Канада. За разпространението на северноамериканските кафяви мечки изключително стратегическо значение има Беринговия проток, който сега отделя Северна Америка от Азия. По време на заледяванията обаче, този проток е представлявал връзка от лед, по която кафявите мечки са навлезли от Азия в Северна Америка и се разпространили там. Това е станало за първи път преди около 70 хил.г. Тази колонизация е съпътствана и с колонизиране и на Тихоокеанските крайбрежни острови. След навлизането си на континента, кафявите мечки се разпространяват на юг и постепенно дивергират генетично, давайки различните сегашни клонове. Втора колонизационна вълна се е осъществила след последния глациален максимум / преди 26-13 хил г/. По време на това заледяване Кордилерите и Лорентидите са били покрити с плътна ледникова покривка и мечките са се изтеглили на юг. Преди около 21 хил год. втора колонизация от мечки, включвайки такива от клон 3в навлизат от Евразия в Берингия. Края на последния глациален максимум е бил преди около 13-9 хил год ,когато ледниците отстъпват постепенно и постепенно Беринговия проток се обособява отново, разделяйки континентите окончателно досега. Малко преди това да стане трета вълна колонизациа на мечки от клон 3а е вече била факт.
Друг интересен аспект на филогенетичното изследване за Северна Америка е положението на полярната мечка. Оказва се, че полярната и кафявата мечка имат парафилетични отношения. Клон 2а на кафявата мечка е изключително близък с клон 2в, който е на полярната мечка, спрямо другите клонове на кафявите мечки. Това показва непълно разделяне на клоновете, поради недостатъчно време за това /от еволюционна гледна точка е минало малко време от разделянето на двата вида мечки/, освен това други изследвания показват, че след разделянето на двата вида е имало хибридизация, която също се е отразила на генофонда. Подобни сложни взаимотношения бяха описани наскоро от учени, които откриват, че мечките от АВС (Admiralty, Baranof and Chichagof (ABC) Islands) острови, които са на външен вид и като поведение кафяви мечки имат митохондриална ДНК, която съответства на тази при полярните мечки. Една от най – вероятните хипотези е, че след затоплянето и разтопяването на леда от последния глациален максимум, полярни мечки са останали изолирани на тези острови. Впоследствие до тези острови преди около 150 хил год. досигт мъжки кафяви мечки, и след няколко поколения се получава сегашния генофонд на тази популация.
За Европа, клон 1 а е разпространен в южна Скандинавия и югозападна Европа – Испания и Франция; клон 1в на юг и изток в Европа – Италия, Словения, Хърватия, Босна, Гърция, България, Румъния, Полша; клон 3а в североизток и изток на Европа – северна Скандинавия, Финландия, Русия , Естония, Словакия и Румъния. Контактна зона между 1 а и 3а има в Швеция, а между 1 в и 3а в Румъния.
Все още не са окончателни данните кога за първи път кафявата мечка се е появила в Европа. Има намерени фосили на смесен вида на кафява и пещерна мечка отпреди около 900 хил год – ранен плейстоцен. Най – вероятния предшественик на съвременната европейска кафява мечка датира от късен плейстоцен / 175-263 хил г/. По време на последния глациален максимум, когато Европа е била покрита с ледници, популациятана кафявите мечки се свива на юг и остава в рефугиуми , при подходящи по – меки условия. За клон 1а – Иберийски полуостров, за клон 1 в – Италийски и Балкански полуострови, за клон 3а не е уточнено местоположението , но се предполага , че е бил в Кавказ и Карпатите. От тези рефугиуми при благоприятни вече условия, когато климата се затопля в началото на холоцена, мечките реколонизират обратно Европа. Генетичното разнообразие е вече значително по-ниско, от гледна точка на ограниченото разпространение по време на заледяването, бързата реколонизация и последващо влияние на човека. Процесите на разпространение на кафявата мечка все още са в процес на изследвания, поради новите данни, които постъпват. Изясняват се следните вероятни събития: мечките са били малко по-разпространени на север по време на глациалния максимум или са имали още един рефугиум разположен в Централна Европа. При изследване на популациите в Скнадинавия като контактна зона се откриват и мечки от клон 1в, като предварително се е очаквали контактна зона между 1 а и 3а. Това води до извода ,че мечките от клон 1в, с рефугиум Итало – Балкански полуострови, са започнали по- ранна експанзия на север, отколкото предварително се е смятало.
Клон 3а е един от най – широко разпространените клонове в цяла Холарктика от Евразия до Аляска, включително част от Япония. Най – вероятния предшественик на този клон е живял преди около 26 хил г. Клон 3в е разпространен в Япония и Аляска , Канада. Клон 4 – Япония, Канада, САЩ.
Клонове 5, 6 и Иранския са с неуточнена позиция. Клон 2в е разпространен циркумполярно и е хаплотипа на полярната мечка.
За клон 1а Иберийски и клон 1в Итало – Балкански е установен сигурен маркер за отличаване – единична мутация на място 16 762 в контролния регион на мтДНК. При клон 1а хаплотипове на това място има база А – аденин, а при хаплотип 1в база G – гуанин.
Природозащита на кафявата мечка в България
В България кафявата мечка се счита, че има две популации – Рило – Родопска и Старопланинска. По последни данни популациите наброяват около 1000 мечки. В Екшън плана за кафявата мечка в Европа, публикуван през 2000 г за България се предписват редица препоръки за опазване, изследване и решаване на проблемните мечки. Една от препоръките е да се предвидят компенсации на пострадалите фермери и хора от населението в контактните зони, с цел да се намали основния проблем, взаимодействие човек – мечка. През 2008 в България се приема План за действие за опазване на кафявата мечка. Определят се работни групи по наблюдение и събиране на материали. Определят се работни региони. Видно е, че средства почти няма. Не се предвижда и решение на проблема с отношението човек – мечка. През 2012 година е приета забрана за отстрела на кафявите мечки в България.
Насоки за опазване на вида кафява мечка в България
оценка на съществуващите Рило-Родопска и Старопланинска популации
неинвазивно събиране на материал за изолиране на ДНК от косми и екскременти.
ДНК изследвания в следните насоки
Типизиране на хаплотиповете – очакван хаплотип Итало-Балкански клон 1в, но е възможна миграция и установяване на източен хаплотип 3а, който може да е от интродукцията на Карпатски мечки и/или от миграция на запад на мечки от Евразия още от последния глациален максимум.
Оценка на генетичния полиморфизъм на съществуващите популации, което да бъде използвано за оценка на жизнеността на популацията – колкото е по-високо нивото на полиморфизъм, толкова по-жизнена е популацията, ниския генетичен полиморфизъм е показател за уязвимостта на популацията
ДНК секвениране и депозиране на информацията в световната ГЕНБАНК
Развиване на гъвкава и бърза система от компенсации на пострадали от мечки фермери, хора от населението в контактните зони и/или други методи , с цел намаляване конфликта човек/мечка
Нормалният сърдечен ритъм е около 40 удара в минута, при зимен сън ритъма пада до 8!
Ноктите на мечките се различават в зависимост от начина им на живот. Черните мечки, които често се катерят имат силни, извити нокти, а гризли, които често ровят земята имат дълги и прави нокти.
Един от най- любопитните факти около биологията на мечките е т. нар. закъсняла имплантация. След като мечката е оплодена зиготата започва развитие, като стига до етап бластоцист и влиза в състояние на латентност. Без да се имплантира бластоциста изчаква няколко месеца, през което време мечката трупа запаси и ако тя е в добро състояние с достатъчно маса, в добро здраве и не е под стрес следва имплантация и по- нантатъшно развитие. Иначе бластоциста се абсорбира в тялото на мечката. Закъснялата имплантация позволява на мечките да се адаптират по- добре към външните условия и раждането да се случи в най- благоприятния момент от годината.
Всъщност мечките са всеядни животни, като при повечето от тях месото е малък процент от менюто. Едиствения истински хищник сред мечките е полярната мечка. Нейния стомах може да побере до 68 кг месо!
Пандата може да изяде до 20 кг бамбук на ден.
Бърнестата мечка се храни най- вече с термити. При тези мечки липсват горните два резеца, което им позволява да изсмукват като с прахосмукачка насекомите от гнездата им.
Полярните мечки имат изключително обоняние- те могат да проследят миризма от 32 км и да надушат мъртав тюлен под 3 м дебел лед.
Бърнестата мечка е единствената, която е активна нощем.
Около 98% от популацията на гризли живее в Аляска.
Всички мечета се раждат без козина… с изключение на полярните и големите панди.
План за действие за кафявата мечка в България. Министерство на околната среда и водите. София 2008.
Петър Генов, Александър Дуцов*Диана Златанова, Даниело Пешев**ПРЕДВАРИТЕЛНО ПРОУЧВАНИЯ ЗАРАЗМНОЖАВАНЕТО НА МЕЧКАТА(URSUS ARCTOS L.) В РОДОПИТЕ. ГОДИШНИК НА ШУМЕНСКИЯ УНИВЕРСИТЕТ “ЕПИСКОП КОНСТАНТИН ПРЕСЛАВСКИ”Т. ХVІІІ В 6 ПРИРОДНИ НАУКИ Факултет по природни науки. Том ХVІІІ В 6, 2008 г
John Davison , Simon Y.W. Ho, Sarah C. Bray , Marju Korsten, Egle Tammeleht, Maris Hindrikson,Kjartan Østbye , Eivind Østbye , Stein-Erik Lauritzen , Jeremy Austin , Alan Cooper , Urmas Saarma.Late-Quaternary biogeographic scenarios for the brown bear (Ursus arctos) a wild mammal model species. Quaternary Science Reviews 30 (2011) 418-430
Bray S.C.E. 2010. Mitochondrial DNA analysis of the evolution and genetic diversity of ancient and extinct bears. Thesis of PhD of University of Adelaide. 256p.
A. COTOVELEA, N. ŞOFLETEA, G. IONESCU, O. IONESCU. GENETIC APPROACHES FOR ROMANIAN BROWN BEAR (Ursus arctos) CONSERVATION. Bulletin of the Transilvania University of Braşov.Series II: Forestry • Wood Industry • Agricultural Food Engineering • Vol. 6 (55) No.1 – 2013
Ce´ line Bon, Nicolas Caudy, Maud de Dieuleveult, Philippe Fosse, Michel Philippe, Fre´de´ ric Maksudd,E´liane Beraud-Colomb, Eric Bouzaid, Rym Kefie, Christelle Laugier, Bernard Rousseau, Didier Casane, Johannes van der Plicht, and Jean-Marc Elalouf.Deciphering the complete mitochondrial genome and phylogeny of the extinct cave bear in the Paleolithic painted cave of Chauvet. PNAS _ November 11, 2008 _ vol. 105 _ no. 45 _ 17447–17452
7. James A. Cahill, Richard E. Green, Tara L. Fulton, Mathias Stiller, Flora Jay, Nikita Ovsyanikov,
Rauf Salamzade, John St. John, Ian Stirling, Montgomery Slatkin, Beth Shapiro. Genomic Evidence for Island Population Conversion Resolves Conflicting Theories of Polar Bear Evolution. PLOS Genetics, March 2013 | Volume 9 | Issue 3 | e1003345
8. Marju Keis.2012. Brown bear /Ursus arctos/ phylogeography in northern Eurasia. Thesis of PHD. University of Tartu. Estonia. 65p.
9. Lisette P.Waits,* Jack Sullivan,† Stephen J. O’Brien,‡ and R. H.Ward. Rapid Radiation Events in the Family Ursidae Indicated by Likelihood Phylogenetic Estimation from Multiple Fragments of mtDNA. Molecular Phylogenetics and Evolution Vol. 13, No. 1, October, pp. 82–92, 1999
10. Marie Page`s, Seґbastien Calvignac, Catherine Klein, Mathilde Paris,Sandrine Hughes, Catherine HaЁnni. Combined analysis of fourteen nuclear genes refines the Ursidae phylogeny. Molecular Phylogenetics and Evolution 47 (2008) 73–83
Книги, които те прегръщат; книги, които те усмихват; книги, които те стоплят. До сега съм писала само за такива екземпляри. Но тази „Марика“ просто се вкопчи в мен, раздруса ме и се изпрати за минути.
Сафон те хваща за ръката и те води по улиците на Барселона. Хватката му е мека, но уверена. Сюжетът е прост, но загадъчен. Героите обникновени, но истински. От онези, чиито чувства стават твои. А Марина… Първо помислих, че е дух, после ангел, накрая тя се превърна и в двете. Но не се подлъгвайте по заглавието, тя не е главното действащо лице.
Неочаквано това е ТОЙ и не е влюбеният в нея младеж. Честно казано Оскар и Марина сами си намират белята, отивайки на „невинна” разходка на гробището, за да срещнат странната забулена в черно жена. Тръгват след нея, а зад тях незабележимо или не толкова, се промъква цяло море от разкрития.
През цялото време, четейки романа, аз се мъчих да го оприлича на нещо. Ту приличаше на „То” с емоционалното напрежение, което създава у теб, ту биеше на „Тайната градина” с романтичната мистерия на двама младежи. Мъчих се, мъчих се, а накрая осъзнах, че този роман няма описание. В него има всичко от любов до смърт, от ужасен страх до романтична нежност. Хвърлях книгата настрана, за да я отворя след 5 минути отново. Криех се под одеялото от нея, а когато четях до 3 часа ме беше страх да изгася лампата.
Но това беше хубавият страх, който гъделичка съзнанието ти, рисува картини пред теб, толкова реални, че вярваш в тях. Сафон е невероятен художник, който създава серия от картини пред теб, с неповторими и леки щрихи. Това е първата книга на този автор, която попадна в ръцете ми. И две мнения няма – тя се превърна в първата от многото бъдещи.
Наука за химичния състав, свойства, обмяна на веществата и енергията в живите организми. Биохимията изучава животът на молекулно ниво. За живота както и за други феномени не е възможно да се даде прецизна дефиниция. Norman Нorowitz предлага следните критерии за живите организми: Животът обхваща свойствата на репликация, катализа и променливост, тези свойства са характерни за почти всички живи организми. Всички биологични процеси са химични трансформации.
БИОГЕННИ ЕЛЕМЕНТИ
От познатите химични елементи 90% от тях са намерени в природата, а по-голямата част от тях се намират в биосферата, но за живата материя са от значение сравнително малък брой химични елементи – C, H, O, N, S, P, Cl, Na, K, Ca, Mg, Zn, Fe, Cu, Co, Mo, J.
Според количественото си съдържание в организмите биогенните елементи се разделят на 2 големи групи: Макро и Микро елементи;
МАКРО ЕЛЕМЕНТИ
Съставляват приблизително 99% от елементите от биосферата 6 от тях играят изключително важна роля за живите организми – C, H, N, O, P, S ; те изграждат всички биологични макромолекули, централно място заема въглерода (С). Сравнително малкия му атомен радиус и междинната му електроотрицателност определя образуването на типичните ковалентни връзки. Не по-малка е и ролята на кислорода (О) той образува единични или двойни връзки поради по-високата си електроотрицателност. Връзките му с въглерода са типично полярни (нуклеофилни заместителни реакции)
Азота (N) също образува ковалентни връзки в органичните съединения. Особено важна роля на неподелената свободна електронна двойка на N. Taзи електронна двойка придава по-голяма или по малка степен основен характер. Органогенните елементи фосфор (Р) и сяра (S) образуват връзки със значително по-ниска здравина. С участие на фосфора се образуват т.нар макроергични съединения (АТФ). А със сярата (ацетил-КоА). Ако може да се твърди, че въглеродните атоми изграждат скелет на всички биогенни молекули то най-малкия органогенен елемент водорода (Н) запълва празнините в този скелет, като образува водородни връзки с по-електроотрицателните елементи кислород (О) и азот (N). Водорода участва при формирането на вторичните и третичните структури на биогенните макромолекули. Освен водата, която е основна среда в организма в състава й участват неорганични съединения под формата на йони – H , OH, H2PO-4, HPO2-4, PO3-4, HCO-3, CO2-3, SO2-4, NH 4;
Освен органогенните елементи, които изграждат цялата структура на клетката останалите елементи предимно са съсредоточени в определени органи. В тези органи те изпълняват специфични функции – Na и K се срещат в организма само като катиони и те образуват йонни връзки за разлика от по-горе споменатите елементи. Докато Na се съдържа в извън клетъчната течност то K се намира в клетката. По-този начин те участват в подържането на осмотично налягане от двете страни на клетъчната мембрана. Калция(Са) и магнезия (Mg) имат склонност към образуване на комплекси съединения, Са участва в клетните образувания в организма и изпълнява специфични функции при контракция на мускулите. Магнезия участва като активен център в много фосфатази.
МИКРО ЕЛЕМЕНТИ
Общото количество на микро елементите в биосферата е около 1%. Забелязано е тясна връзка между заниженото или повишено съдържание на даден микроелемент в определен орган и появата на характерно заболяване. Например намаленото съдържание на йод в щитовидната жлеза води до болестта „Гуша” и Кретенизъм – умствена недостатъчност. Действието на микроелементите е в тясна свръзка с това на хормоните, ензимите и витамините т.е микро елементите участват преди всичко в регулаторните механизми на организма. Особено важно е участието на желязото – характерна структурна единица Fe – в профинилов комплекс. Този комплекс участва в изграждането на хемоглобина – приносител на кислород или в цитохромни системи, участващ в окислително-редукционни комплекси. Съдържанието на желязо е около 5g на 70kg тегло, 70% участва в хемоглобина а останалата част(30%) е разпределена предимно в черния дроб, бъбреците и кръвния серум. Общото съдържание на цинк (Zn) e около 3g за 70kg тегло. Богати на цинк са жлезите с вътрешна секреция особено половите!, черния дроб и тъканите на окото. Цинка е активен център в ензимите: Алкохолдехидрогеназа, Карбоксилпентидаза и Карбоанхидраза. Богати на мед (Cu) са черният дроб надбъбречна жлеза и сивото вещество на главния мозък. Медта лесно преминава от първа във втора валентност и обратното и по тази причина тя участва в редокси процесите.
Мангана (Mn) влиза в състава на различните ензими и предимно той изпълнява активаторна функция. Съдържанието на кобалт (Со) е твърде ниско, богати на кобалт са жлезите с вътрешна секреция и кърмата. Той е съставна част от витамин В12. Редица заболявания като анемия, хепатит са свързани с нарушената обмяна на кобалта в организма. Някой елементи в много-ниска концентрация проявяват токсично действие!
ТОКСИЧНИ ЕЛЕМЕНТИ
Алуминий(Al) – Причинява анемия тъй като измества желязото от неговите съединения. Той измества и калия от съединенията му като образува по-стабилни връзки със същите лиганди.
Арсен(As) – Арсен в пета валентност симулира фосфор в пета валентност. Арсена взаимодейства с тиолите или силго групите от протеините и инхибира ензимните системи.
Берилий(Be) – Най-токсичния елемент! Измества магнезия от всичките му съединения. Свързва се много здраво с нуклеиновите киселини.
Кадмий(Cd) – Измества цинка от всичките му съединения.
Живак(Hg) – Акумулира се от черния дроб, бъбреците и мозъка, нарушава техните функции.
Олово(Pb) – Образува здрави връзки в –SH групите и инхибира ензимите.
Стронций(Sr) – Симулира калция
Талий(Tl) – Нервотоксичен! Стабилен йон, измества калия от съединенията му и нарушава биологичните функции.
Американски учени се доближават до момента, в който ще съумеят да създадат съвсем ново възобновяемо гориво
Учените са с една стъпка по-близо до създаването на възобновяемо гориво, близко до петрола, в чийто състав са включени някои бактерии, въглеродният диоксид и слънчевата светлина.
Джанис Фриас, докторант по инженерна химия в Университета в Минесота открила как да използват протеини, за да превърне мастни киселини, произведени от бактерии, в кетони, които могат да бъдат преобразувани така, че да се създаде въглеводородно гориво. Адитя Бхан и Лани Шмид са включени в нейния екип.
Професорът по биохимия Лари Уакет споделя: „Въглеродният диоксид е основният парников газ, водещ до глобалното изменение на климата, така че да се маха от атмосферата по някакъв начин е добро за околната среда. Това е също така и безплатно. И скоро ще можем да използваме една и съща инфраструктура за обработката и транспорта както на изкопаемите горива, така – и на този нов вид гориво, произведено от въглеводород.“
Изследователите използват Synechococcus – бактерия, която определя съдържанието на въглеродния диоксид в слънчевата светлина и го преобразува на захари, които след това се дават на Shewanella – друга бактерия, която произвежда въглеводороди, които след това се превръщат в парникови газове.
Въглеводородите (изработени от въглерод и водород) са основният компонент на изкопаемите горива. За производството на изкопаемите горива на Земята са й необходими стотици милиони години, а човечеството ще съумее да изчерпа запаса от тях само след около 50 години.
Учените са финансирани от американското Министерство на енергетиката по програма за финансиране на перспективни научни изследвания.
Изследователи от Вашингтонския Университет (UW) и Университета на Северна Каролина в Чапъл Хил (UNC) са направили важна стъпка за превръщането на метана от газ в течност, като това го прави по-използваем като гориво и източник за производство на други химични вещества.
Метанът, основният компонент на природния газ, се намира в изобилие и е атрактивен като гориво и суровина за химикали, защото е по-ефикасен от нефта, причинява по-малки замърсявания и може да служи за заместител на петролните горива докато се достигне широко разпространение на възобновяемите горива.
Транспортирането на метан обаче е трудно и скъпо, защото той остава в газообразно състояние при температури и налягане типични за земната повърхност
Сега учените от UNC и UW се придвижиха по-близо към начин да се преобразува метана в метанол или други течности, които могат лесно да бъдат транспортирани, включително и от по-отдалечени места, където метанът е често откриван. Откритието е публикувано в Science, броя от 23 октомври.
Метанът е ценен заради своите високоенергийни въглеродо-водородни връзки, които се състоят от въглероден атом свързан с четири водородни атома. Газът не влиза лесно в реакция с други материали и най-често просто е изгарян в качеството си на гориво. Горенето предизвиква разпадане на всички въглеродо-водородни връзки и в резултат се получава въглероден двуокис и вода, казва Карен Голдбърг, професор по химия в UW.
Преобразуването на метана в полезни химични вещества, включително транспортируеми течности понастоящем изисква високи температури и много енергия. Открити са катализатори, които превръщат метана в други химични вещества при ниски температури, но те са твърда бавни, твърде неефективни или прекалено скъпи за промишлени цели, казва Годлдбърг.
Въздействието върху метана с метални катализатори е първата стъпка необходима са селективно премахване на една от въглеродо-водородните връзки в процеса на преобразуване на газа в метанол или друга течност. В статията си, изследователите описват първото наблюдение на метален комплекс (съединение състоящо се от централен метален атом свързан с окръжаващи го атоми или молекули), който преобразува метана в течност. Това съединение служи за модел за други възможни метанови комплекси. В тези комплекси, метановите въглеродо-водородни връзки остават непокътнати, като се свързват с рядък метал, наречен родий.
Изследването трябва да даде тласък на по-нататъшно развитие на катализатори, под чието въздействие метанът се превръща в метанол или други течности, казва Голдбърг, въпреки че тя отбелязва, че действителната разработка на процеса до степен превръщане на газа в течност при приемливи температури е вероятно да стане някога в бъдещето.
„Идеята е да се преобразува метанът в течност, като при това се запазят повечето въглеродо-водородни връзки, така че да се запази цялата енергия”казва тя. „Това ни дава следа как трябва да изглежда първото взаимодействие между метана и метала”
Морис Брукхарт, професор по химия в UNC, казва, че въглеродо-водородните връзки са много здрави и е трудно да бъдат разрушени, но разрушаването е по-лесно при метановите комплекси.
„Следващата стъпка е да използваме познанието придобито от това откритие за да формулираме други комплекси и условия, които ще ни позволят каталитично да заменим един водороден атом от метана с други атоми и да създадем течни химични вещества като метанола” казва Брукхарт
Водещ автор на публикацията е Уесли Бърнскотър от Университета Браун, който е участвал в изследването, докато е работил в UNC. Голдбърг, Брукхарт и доцент Синтия Шауър от UNC са съ-автори.
Изследването е извършено в лаборатория финансирана от Националната научна фондация – базирания в UW Център за осъществяване на нови технологии чрез катализа, който включва 13 университета и изследователски центрове САЩ и Канада. Допълнително финансиране е осигурено от Националните здравни институти на САЩ.
Центърът ръководен от Голдбърг има за цел откриването на ефикасни, сравнително евтини и не нанасящи вреда на околната среда начини за производство на химикали и горива.
По материали предоставени от Вашингтонския Университет
Кварцът е ключово парче от сложния пъзел, свързан с разбирането на земетресенията
С подземните натрупвания на кварц по света могат да се обяснят земетресенията, формирането на планините и континенталната тектоника, според резултатите от изследване на американски и британски специалисти.
Това откритие може да подпомогне предсказването на земетресенията и да отговори на въпросите, свързани с формирането и местонахождението на земетръсните разломи, планините, долините и равнините.
„Въпросът защо планините са се образували там, където са, е зададен в дълбините на времето. Той е стар почти колкото света.“, казва геофизикът Антъни Лаури от университета на Юта, един от авторите на проучването.
Учените изследвали температурата и гравитацията в западните щати на САЩ със специална апаратура, за да опишат геоложките свойства на земната кора.
Те установили, че в щати като Калифорния, Айдахо, Невада и Юта има натрупвания на кварцови кристали на местата, където се намират планините и линиите на разломите. С помощта на специална технология, известна като Earthscope, учените са открили, че кварцът е сигнал за слабост в земната кора, в резултат на която може да се случи геоложко събитие като земетресение или изригване на вулкан.
С кварца може да се обясни и движението на континентите, известен с термина „континентален дрейф” и тектониката на плочите.
Например, мощното земетресение в Япония на 11.03.2011 г. е приближило островната държава с осем фута (или – с ок. 250 см.) до континенталната част на САЩ. Това е станало в резултат на приплъзването на Азиатската тектонска плоча под Северноамериканската тектонска плоча.
Специалистите откриват връзка между свойствата на кварца и земното движение. Съдържащата се в кварца вода се освобождава, когато се загрее под натиск и позволява на скалите да се приплъзват.
Докато за региони като Япония, Южна Калифорния и националния парк Йелоустоун се знае, че са в активната фаза на т. нар. вискозен цикъл, за други региони като Апалачите в източната част на Съединените щати се предполага, че са в неактивна фаза.
Теорията на Лаури и колегите му може да се приложи при оценяването на потенциалния риск от земетресения и тяхната сила, както – и за избирането на безопасни места за изграждане на атомни електроцентрали и големи язовири.
Учените предполагат, че земетресение е било виновно и за преобръщането на течението на р. Мисисипи през 1812 г.
Уилард Ф. Либи (1908 – 1980) е американски химик от екипа, създал атомната бомба. Другият му известен принос е радиовъглеродният метод за установяване на възрастта на археологичните останки. Въглерод 14 съществува в живите тъкани в определено съотношение и се губи с постоянна скорост. Докато организмът е жив, неговото съдържание се поддържа благодарение на метаболизма. След смъртта започва намаляване на количеството му. Либи е изчислил как да се определи времето, минало от смъртта, чрез оценка на количеството въглерод 14, който все още се съдържа в останките. През 1960 г. Либи е награден с Нобеловата награда за химия.
Дезоксирибонуклеиновата киселина (ДНК) е открита през 1929 г. от Фебъс Арън Теодор Левен (1869 -1940), руски учен, работещ в Америка. Уникалната двойна спирална структура на молекулите на ДНК е описана през 1953 г. в Англия като резултат на обединените усилия на два екипа от изследователи. През тази година Франсис Крик (роден 1918) и Джеймс Уотсън от Америка (роден 1928), работещи в Кембридж, създават първия модел на молекулярната структура на ДНК, който отговаря на всички експериментални характеристики на субстанцията. Тяхната работа става възможна благодарение на откритията и идеите на Розалин Франклин (1920 – 1958) и Морис Уилкинс от Нова Зеландия (роден 1916), които работят заедно близо до Лондон и изучават молекулата на ДНК. През 1962 г. тримата, които са останали живи, си поделят Нобеловата награда за това откритие. Преди да е намерена неговата структура, вече са били разкрити наследствените функции на ДНК.
Дезоксирибонуклеиновата киселина (ДНК) е нуклеинова киселина, която носи генетичните инструкции за биологическото развитие на всички клетъчни форми на живот и много от вирусите. ДНК понякога се нарича молекула на наследствеността, тъй като тя се наследява и се използва за разпространение на признаци. При възпроизводството тя се реплицира и се предава на потомството.
При бактериите и други прости клетъчни организми ДНК е разположена някъде из цялата клетка. В сложните клетки, от които са изградени растенията, животните и в други многоклетъчни организми, повечето ДНК ев хромозомите, които се намират в клетъчното ядро. Органелите–енергийни генератори, познати като хлоропласти и митохондрии също носят ДНК, както и много вируси.
* Гените могат да се разглеждат грубо като готварска книга на организма.
* Една верига ДНК съдържа гени – области, които регулират гените – и други области, които нямат функции или функциите им все още не са познати;
* ДНК се състои от две нишки с връзки помежду им, които могат да се разделят подобно на цип;
* ДНК кодира генетичната информация благодарение на четири „строителни елемента”, наречени бази: аденин, тимин, гуанин, цитозин. Те се обозначават съкратено като А, Т, Г и Ц и имат свойството всяка да „се чифтосва” само с една от останалите три бази: А Т, Т А, Г Ц, Ц Г; такаче едно „А” на едната нишка от ДНК ще се „съеши” успешно само с „Т” от другата нишка.
* Редът е от значение: А Т не е еквивалентно на Т А, както и Ц Г не е едно и също с Г Ц.
* Но, тъй като са възможни само 4 комбинации, базите на едната от нишките са достатъчни, за да се опише последователността.
* Редът, в който са разположени базите по дължината на ДНК, е важен —ДНК-последователността (или секвенцията) е описанието на гените.
* Репликацията или удвояването на ДНК се осъществява чрез разделяне на двете нишки с относително прости химически реакции и създаване на„втората половина” на така получената единична верига чрез потапяне в„супа”, която съдържа всичките четири бази. Тъй като всяка база може дасе комбинира само с една от останалите три бази, подредбата на базите в съществуващата верига определят еднозначно какви бази ще има в новообразуваната верига и как ще са подредени. По този начин, всяка единична верига образува точно копие на оригиналната ДНК, като събира необходимите бази в „супата”, освен ако не настъпи мутация.
* Мутациите са просто химически грешки в този процес: една база може случайно да бъде пропусната, вмъкната или грешно копирана или пък веригата може да бъде скъсена или удължена; всички други основни мутации могат да се опишат като комбинация от тези случайни „операции”.
ДНК е позната на учените от повече от сто години. Първоначално е идентифицирана през 1868 г. от швейцарския биолог Фридрих Мишер в сперматозоиди от сьомга. Той нарича откритата от него субстанция нуклеин, но въпреки това ДНК е призната официално едва през 1943 г. в резултат на експеримент, реализиран от Осуалд Ейвъри. Структурата на двойната ДНК спирала е била открита през 1953 г. от Джеймс Уотсън и Франсис Крик в Кеймбриджкия университет. За това си откритие те получават Нобелова награда за химия.
http://bgnauka.eu/pic/bio/DNA1.jpg
Въпреки че понякога ДНК е наричана „молекулата на наследствеността”, парчетата ДНК, както си ги представят повечето хора, не са единични молекули. По-скоро те са двойки молекули, които се преплитат като лиани,образувайки двойна спирала.
Всяка лианообразна молекула е ДНК верига: химически свързана верига отнуклеотиди, всеки от които се състои от захар, фосфат и един отчетирите вида ароматно-въглеводородни „бази”. Тъй като нишките в ДНА са изградени от тези нуклеотидни елементи, те са полимери.
Тъй като има четири вида бази, има и четири вида нуклеотиди, които често се обозначават като своите бази. Тези бази са аденин (А), тимин (Т), цитозин (Ц) и гуанин (Г).
Полинуклеотидните вериги в двойната верига на ДНК могат да се свързват чрез хидрофобния ефект. Това, кои вериги остават свързани, се определя от комплементарното свързване. Всяка база образува водородни връзки само с една от останалите бази — А с Т, Ц с Г — така че видът на базитена едната верига определя силата на връзката — колкото повече комплементарни бази съществуват, толкова по-силна и устойчива е връзката.
Механизмите в клетката могат да разделят двойната спирала на ДНК и такавсяка единична верига може да служи за шаблон за създаване на нова верига, почти еднаква с предишната. Грешките при синтеза се наричат мутации. Процес, наречен ПВР имитира този процес ин витро в не жива среда.
Тъй като при свързване в двойки нуклеотидните бази се обръщат към остана спиралата, а захарните и фосфатните групи са от външната страна,двете вериги, които те образуват, изглеждат като носещи елементи на спиралата. В действителност химическите връзки между фосфатите и захарите свързват един нуклеотид със съседните му във веригата.
Последователността от нуклеотиди по една ДНК-верига в един ген дефинира един белтък, който организмът трябва да бъде произведе или както е прието да се казва, да „експресира” в един или няколко момента отживота си, като използва информацията, съдържаща се в последователността. Отношението между нуклеотидната последователност и тази от аминокиселини в белтъка се определя от прости клетъчни правилана транслация, познати под името генетичен код. Генетичният код се състои от трибуквени думи (наречени кодони), образувани от последователност от три нуклеотида (напр. АЦТ, ЦАГ, ТТТ). Тези кодони след това могат да се транслират посредством информационна рибонуклеинова киселина (РНК) и впоследствие транспортна РНК до кодон,съответстващ на определена аминокиселина. Тъй като има 64 възможни кодона, повечето аминикиселини имат повече от един възможен съответе нкодон. Има също така три „стоп” или „безсмислени” кодо на, озачаващи края на кодовата последователност.
Изглежда, че при много видове организми само малка част от цялата последователност на геномa кодира белтъци. Функцията на останалата час те предмет на спекулации. Знае се, че някои нуклеотидни секвенции задават афинитет към ДНК свързващи белтъци, които играят различни жизненоважни роли, по-специално чрез контрол на репликацията и транскрипцията. Тези последователности често се наричат регулационни секвенции и изследователите предполагат, че засега са идентифицирали само една малка част от съществуващите. „Отпадъчната ДНК” представлява участъци, които не изглежда да съдържат гени или да имат някаква функция.
Секвенцията определя и податливостта на даден сегмент от ДНК на разкъсване, причинено специфично от ензими – рестрикционни ендонуклеази (съкр. рестриктази)— най-типичното средство на генното инженерство.Позициите на местата на разкъсване след третиране на генома на даден индивид с определен ензим са строго специфични. В практиката се използва третиране само на определен участък, при което може да се състави ДНК профил на индивида (един от множеството възможни).
Репликация или синтез на ДНК е процесът на копиране на двойната верига на ДНК преди деление на клетката. Двете получени двойни вериги са почти съвсем еднакви, но понякога грешки при репликацията могат да доведат дополучаване на не съвсем точно копие (виж мутация) и всяка от тях се състои от една оригинална и една ново синтезирана верига. Това се нарича полуконсервативна репликация. Процесът на репликация се състои от три стъпки: инициация, репликация и терминация.
Водородните връзки между веригите на двойната спирала са достатъчно слаби, за да могат лесно да бъдат прекъснати с помощта на ензими. Ензими, познати като хеликази разсукват веригите, за да подпомогнат действието на ензими, четящи последователността като ДНК-полимеразата. При разсукването ензимите трябва да прекъснат по химически на чинфосфатният гръбнак на едната от веригите, за да може тя да се завърти около другата верига. Веригите могат също така да бъдат разделени чрез леко нагряване, използвано например в ПВР, ако имат по-малко от около10 000 базисни двойки (10 кило базисни двойки или 10 кбд). Усукването на ДНК-веригите затруднява разделянето на дълги сегменти.
Когато двата края на една двойна спирала на ДНК са съединени, така че тя образува кръг, както е при плазмидите, веригите са топологично заплетена. Това означава, че те не могат да бъдат разделени чрез леко нагряване или друг процес, който не включва прекъсване на едната верига. Задачата за разплитане на топологично свързани вериги на ДНК се изпълнява от ензими, наречени топоизомерази. Някои от тези ензими разплитат цикличната ДНК чрез прекъсване на двете вериги, така че друг двойно верижен сегмент да може да мине между тях. Разплитането е необходимо за репликацията на циклични ДНК, както и за различни видове рекомбинация в линейни ДНК.
Спиралата на ДНК може да приеме една от три леко различаващи се форми.За преобладаваща в клетките се счита „B” формата, описана от Джеймс Уотсън и Франсис Крик. Тя е широка 2 на нометра и има дължина 3,4нанометра на 10 базисни двойки (бд) от последователността. Средната дължина на стъпката на спиралата — последователността, в продължение на която спиралата прави едно пълно завъртане около оста си – е също толкова. Честотата на завъртанията зависи до голяма степен от притеглящите сили, които всяка база упражнява върху съседите си във веригата.
Малката ширина на двойната спирала прави намирането й чрез конвенционална електронна микроскопия невъзможно, освен чрез интензивно оцветяване. В същото време ДНК в много клетки може да бъде с макроскопична дължина — около 5 сантиметра за веригите в една човешка хромозома. Следователно клетките трябва да сбиват или „пакетират” ДНК,за да могат да я побират. Това е една от функциите на хромозомите,които съдържат белтъци, подобни на макари, наречени хистони, околокоито ДНК се навива.
При липса на напрежение B-формата на ДНК-спиралата се усуква на 360° на всеки 10,6 базови двойки. Но много процеси в молекулярната биология могат да причинят напрежения. Един сегмент от ДНК с недостатъчно или прекалено усукване на спиралата се нарича съответно положително или отрицателно „пренавит”. ДНК ин виво обикновено е отрицателно пренавита,което подпомага разсукването на двойната спирала, необходимо за РНКтранскрипция.
Другите две познати двойно-спирални форми на ДНК, наречени A и Z, се различават донякъде по своята геометрия и размери. Изглежда възможно A-формата да се среща само в дехидрирани проби от ДНК като тези,използвани в кристалографски експерименти, и може би в хибридни свързвания на ДНК и РНК-вериги. ДНК-сегменти, които клетките са метилирали с регулиращи цели, могат да възприемат Z-геометрия, при която веригите са усукани около оста на спиралата като огледален образна B-формата.
Поради несиметричната форма и свързване на нуклеотидите една ДНК-верига винаги има видима ориентация. Поради тази ориентация внимателното разглеждане на една двойна спирала показва, че нуклеотидите са ориентирани в една посока по едната верига („възходящата верига”) и в обратната посока по другата верига („низходящата верига”). Такова разполагане на веригите се нарича анти паралелно.
Заради химическата номенклатура хората, които работят с ДНК, наричат асиметричните краища на едната верига 5′ и 3′ краища(произнасят се „пет прим” и „три прим”). Работещите с ДНК, (а също така и ензимите), винаги четат нуклеотидните последователности в „5′ към 3′ посока”. В една вертикално ориентирана двойна спирала 3′ веригата се нарича възходяща, а 5′ веригата — низходяща.
Като резултат от тяхното анти паралелно разположение и предпочитанията на ензимите при четене на последователността, дори и двете вериги да носеха идентични, вместо допълващи се поредици, клетките могат датранслират правилно само едната от тях. Другата верига може да бъдепрочетена от клетката само отзад напред. Молекулярните биолози наричат една последователност „сенс”,ако е транслирана или транслируема, а нейната допълнителна (комплементарна) — „антисенс”. Следователно, донякъде парадоксално,шаблонът за транскрипция е антисенс-веригата. Полученият запис е РНК-копие на сенс-веригата и самият той е сенс.
При някои вируси разликата между сенс и антисенс е размита, тъй като определени секвенции от техните геноми вършат двойна работа, кодирайки един белтък, ако бъдат четени 5′ към 3′ по едната верига и втори белтък, когато се четат в обратна посока по другата верига. Така геномите на тези вируси са необикновено компактни за броя гени, които съдържат. Това се разглежда от биолозите като адаптация.
Тополозите обичат да отбелязват, че разполагането на 3′-края на едната верига на ДНК до 5′-края на другата в двата края на един двойно спираленсегмент прави конструкцията „раков канон”.
В някои вируси ДНК е в неспирална, едноверижна форма. Тъй като много от механизмите за поправка на ДНК в клетките работят само със свързани бази, вирусите, които носят геномите си в едно верижна ДНК, мутиратсравнително по-често. В резултат такива видове могат да се адаптират по-бързо, за да не изчезнат като вид. Резултатът обаче не би бил толкова благоприятен при по-сложни и по-бавно размножаващи се организми, което може да обясни защо само вирусите носят едноверижни ДНК. Тези вируси печелят и от по-ниската „цена“ на репликацията на една верига вместо две.
Полоният, първият открит радиоактивен елемент, е наречен от своята откривателка Мария Кюри (1867 – 1934) на името на нейната родина Полша. Мария Кюри и съпругът й Пиер (1859 – 1906) са извлекли полония от минерала уранит в Париж през 1898 г.
По-късно през същата година Мария Кюри открива по-радиоактивния елемент, радия.
Този сайт използва ‘бисквитки’ (cookies), за да ви предостави възможно най-добро потребителско изживяване. Можете да промените настройките си за бисквитки, или в противен случай приемаме, че сте съгласни с нашите условия за ползване.ПриемамПрочети повече
Правила на поверителност
Privacy Overview
This website uses cookies to improve your experience while you navigate through the website. Out of these cookies, the cookies that are categorized as necessary are stored on your browser as they are essential for the working of basic functionalities of the website. We also use third-party cookies that help us analyze and understand how you use this website. These cookies will be stored in your browser only with your consent. You also have the option to opt-out of these cookies. But opting out of some of these cookies may have an effect on your browsing experience.
Necessary cookies are absolutely essential for the website to function properly. This category only includes cookies that ensures basic functionalities and security features of the website. These cookies do not store any personal information.
Any cookies that may not be particularly necessary for the website to function and is used specifically to collect user personal data via analytics, ads, other embedded contents are termed as non-necessary cookies. It is mandatory to procure user consent prior to running these cookies on your website.


 М16
М16 М16
М16 Слънце
Слънце Червен гигант
Червен гигант Свръхнова
Свръхнова Бели джуджета
Бели джуджета Неутронна звезда
Неутронна звезда Черна дупка
Черна дупка Защо ни впечатляват?
Защо ни впечатляват? За изчезването на видовете
За изчезването на видовете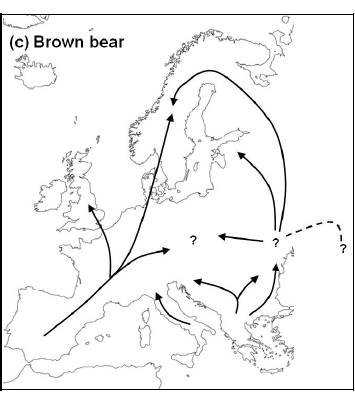 Филогеографските изследвания на основа на митохондриална ДНК дават възможност да се опише начина на разпространение на видовете по време на глациалните цикли. На схемата е показано филогеографското разпространение на кафявата мечка след последния глациален максимум. По време на заледяването се обособяват три рефугиума – Иберийски, Итало-Балкански и източен евразийски ,който е с неуточнено местоположение. Иберийския и Итало-Балканския са били ситуирани съответно на Пиринейския / Иберийски/ , Апенински и Балканския полуострови. От тях при климатичното затопляне започва разселването на вида в Европа и Азия на север, както е показано.
Филогеографските изследвания на основа на митохондриална ДНК дават възможност да се опише начина на разпространение на видовете по време на глациалните цикли. На схемата е показано филогеографското разпространение на кафявата мечка след последния глациален максимум. По време на заледяването се обособяват три рефугиума – Иберийски, Итало-Балкански и източен евразийски ,който е с неуточнено местоположение. Иберийския и Итало-Балканския са били ситуирани съответно на Пиринейския / Иберийски/ , Апенински и Балканския полуострови. От тях при климатичното затопляне започва разселването на вида в Европа и Азия на север, както е показано.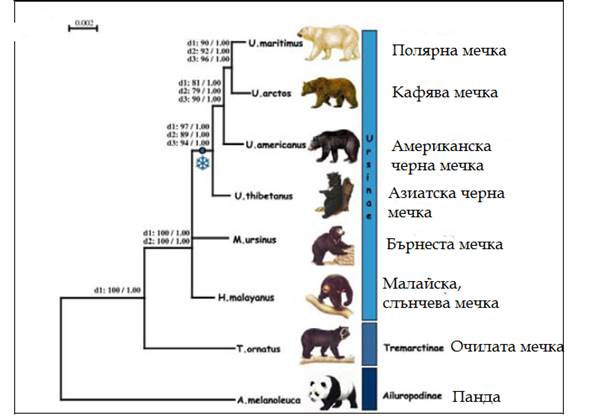
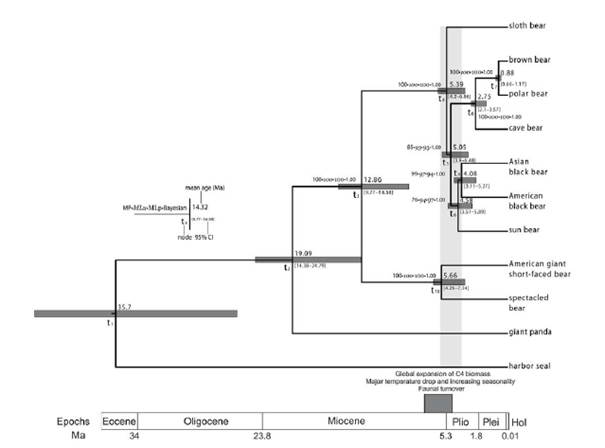 Фиг. 1 Филогения на сем Ursidae, получена на основа на секвениране на региони от мтДНК на съвременни и фосилни видове. Включено е дивергентното време в млн.год.
Фиг. 1 Филогения на сем Ursidae, получена на основа на секвениране на региони от мтДНК на съвременни и фосилни видове. Включено е дивергентното време в млн.год.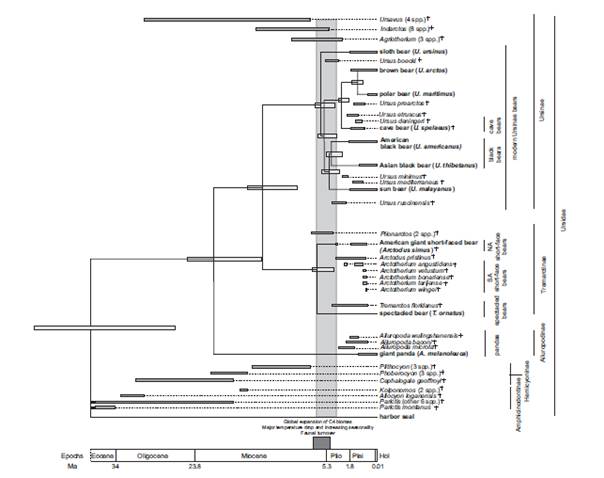







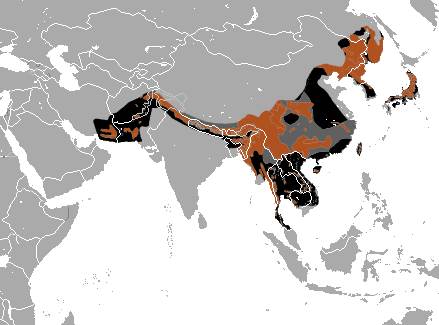






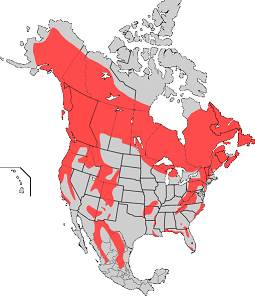






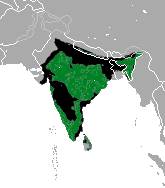 Тази неголяма мечка е уникална сред своите родственици, поради основното им меню насекоми – мравки, термити и др. Бърнестата мечка обитава горските и тревисти райони на Индийския полуостров. Има дълга, пухеста козина, с удължени около рамената участъци, оцветена в черно. Рядко се срещат в канелен, червеникав цвят на козината. Отпред на гърдите имат бяло или кремаво петно в U’ или Y’ форма. Характерна е източната муцуна в по-блед цвят. Формата на муцуната, голите бърни и липсата на горни резци са адаптация за хранене с насекоми. Предните лапи са извити навътре и имат дълги, леко закривени нокти, предназначени за ровене и копаене.
Тази неголяма мечка е уникална сред своите родственици, поради основното им меню насекоми – мравки, термити и др. Бърнестата мечка обитава горските и тревисти райони на Индийския полуостров. Има дълга, пухеста козина, с удължени около рамената участъци, оцветена в черно. Рядко се срещат в канелен, червеникав цвят на козината. Отпред на гърдите имат бяло или кремаво петно в U’ или Y’ форма. Характерна е източната муцуна в по-блед цвят. Формата на муцуната, голите бърни и липсата на горни резци са адаптация за хранене с насекоми. Предните лапи са извити навътре и имат дълги, леко закривени нокти, предназначени за ровене и копаене.









 Храни се основно с плодове и филизи, които събира от дърветата или от земята. Менюто включва допълнително мед, насекоми, малки бозайници, влечуги и рядко добитък. От културните растения предпочитат захарна тръстика, царевица.
Храни се основно с плодове и филизи, които събира от дърветата или от земята. Менюто включва допълнително мед, насекоми, малки бозайници, влечуги и рядко добитък. От културните растения предпочитат захарна тръстика, царевица.





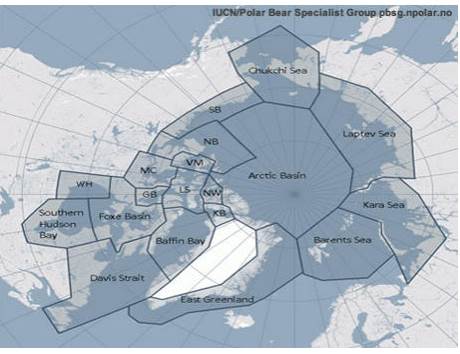 Картата показва разпространението на 19 субпопулации полярна мечка.
Картата показва разпространението на 19 субпопулации полярна мечка.
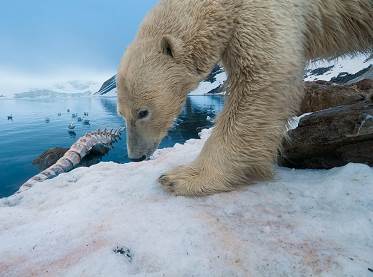


 Наричана на староанглийски още бруун или бруин, което също означава кафява, в Дивия Запад, мечките гризли ги наричали Олд Ефрейм /“Old Ephraim“ / или Мокасиновия Джо /“Moccasin Joe“/, латинското наименование идва от латинската дума мечка – ursus и старогръцката дума мечка – arctos. Кафявите мечки са широко разпространени в голяма част от Северна Америка и Евразия в гористи и планински региони. Този вид е национално животно на много щати в Северна Америка и страни в Европа и Азия.
Наричана на староанглийски още бруун или бруин, което също означава кафява, в Дивия Запад, мечките гризли ги наричали Олд Ефрейм /“Old Ephraim“ / или Мокасиновия Джо /“Moccasin Joe“/, латинското наименование идва от латинската дума мечка – ursus и старогръцката дума мечка – arctos. Кафявите мечки са широко разпространени в голяма част от Северна Америка и Евразия в гористи и планински региони. Този вид е национално животно на много щати в Северна Америка и страни в Европа и Азия.






.jpg)






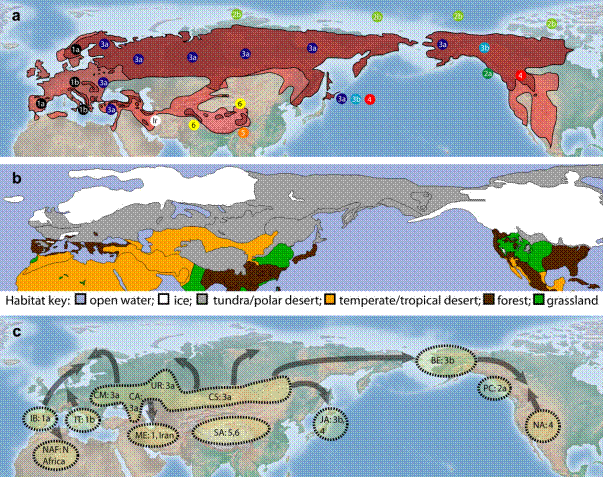












 Наука за химичния състав, свойства, обмяна на веществата и енергията в живите организми. Биохимията изучава животът на молекулно ниво. За живота както и за други феномени не е възможно да се даде прецизна дефиниция. Norman Нorowitz предлага следните критерии за живите организми: Животът обхваща свойствата на репликация, катализа и променливост, тези свойства са характерни за почти всички живи организми. Всички биологични процеси са химични трансформации.
Наука за химичния състав, свойства, обмяна на веществата и енергията в живите организми. Биохимията изучава животът на молекулно ниво. За живота както и за други феномени не е възможно да се даде прецизна дефиниция. Norman Нorowitz предлага следните критерии за живите организми: Животът обхваща свойствата на репликация, катализа и променливост, тези свойства са характерни за почти всички живи организми. Всички биологични процеси са химични трансформации.
 Учените са с една стъпка по-близо до създаването на възобновяемо гориво, близко до петрола, в чийто състав са включени някои бактерии, въглеродният диоксид и слънчевата светлина.
Учените са с една стъпка по-близо до създаването на възобновяемо гориво, близко до петрола, в чийто състав са включени някои бактерии, въглеродният диоксид и слънчевата светлина.
 Изследователи от Вашингтонския Университет (UW) и Университета на Северна Каролина в Чапъл Хил (UNC) са направили важна стъпка за превръщането на метана от газ в течност, като това го прави по-използваем като гориво и източник за производство на други химични вещества.
Изследователи от Вашингтонския Университет (UW) и Университета на Северна Каролина в Чапъл Хил (UNC) са направили важна стъпка за превръщането на метана от газ в течност, като това го прави по-използваем като гориво и източник за производство на други химични вещества.
 С подземните натрупвания на кварц по света могат да се обяснят земетресенията, формирането на планините и континенталната тектоника, според резултатите от изследване на американски и британски специалисти.
С подземните натрупвания на кварц по света могат да се обяснят земетресенията, формирането на планините и континенталната тектоника, според резултатите от изследване на американски и британски специалисти.


 Дезоксирибонуклеиновата киселина (ДНК) е нуклеинова киселина, която носи генетичните инструкции за биологическото развитие на всички клетъчни форми на живот и много от вирусите. ДНК понякога се нарича молекула на наследствеността, тъй като тя се наследява и се използва за разпространение на признаци. При възпроизводството тя се реплицира и се предава на потомството.
Дезоксирибонуклеиновата киселина (ДНК) е нуклеинова киселина, която носи генетичните инструкции за биологическото развитие на всички клетъчни форми на живот и много от вирусите. ДНК понякога се нарича молекула на наследствеността, тъй като тя се наследява и се използва за разпространение на признаци. При възпроизводството тя се реплицира и се предава на потомството.
